Abbreviated Solutions to Problems
Fuller solutions to all chapter problems are published in the Absolute Ultimate Guide to Lehninger Principles of Biochemistry. For all numerical problems, answers are expressed with the correct number of significant figures.
Chapter 1
1.
Diameter of magnified
36,000 mitochondria
glucose molecules
2.
10%
5%
1.6 mm; 800 times longer than the cell; DNA must be tightly coiled
3. Collect the supernatant from the high-speed centrifugation and centrifuge at a very high speed (150,000 g) for 3 hours. The ribosomes will be in the pellet.
4.
Metabolic rate is limited by diffusion, which is limited by surface area.
for the bacterium
Surface-to-volume ratio 300 times higher in the bacterium.
5. (about 23 days)
6. The vitamin molecules from the two sources are identical; the body cannot distinguish the source; only associated impurities might vary with the source.
7.
, carboxyl; , amino; OH, hydroxyl; , methyl
-
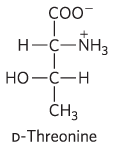
2
8.
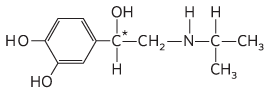
The two enantiomers have different interactions with a chiral biological “receptor” (a protein).
9.
Fatty acids are more nonpolar than amino acids, which makes them separable on the basis of solubility. The larger size and longer shape of fatty acids could allow separation by some types of chromatography.
The charge on the phosphate groups of nucleotides could be used to separate them from glucose. The larger size and shape could allow separation by some types of chromatography.
10. Carbon atoms can form linear chains, branched chains, and cyclic structures. It is improbable that silicon could serve as the central organizing element for life, especially in an -containing atmosphere such as that of Earth. Long chains of silicon atoms are not readily synthesized; the polymeric macromolecules necessary for more complex functions would not readily form. Oxygen disrupts bonds between silicon atoms, and silicon-oxygen bonds are extremely stable and difficult to break, preventing the breaking and making of bonds that are essential to life processes.
11.
(R)-enantiomer: A is ; B is H; C is . (S)-enantiomer: A is ; B is H; C is .
It is unnecessary to make enantiomerically pure (S)-ibuprofen available because the isomerase converts the less effective enantiomer to the effective enantiomer, but does not catalyze the reverse reaction.
12.
3 Phosphoric acid groups; -d-ribose; guanine
Tyrosine; 2 glycines; phenylalanine; methionine
Choline; phosphoric acid; glycerol; oleic acid; palmitic acid
13.
;
-
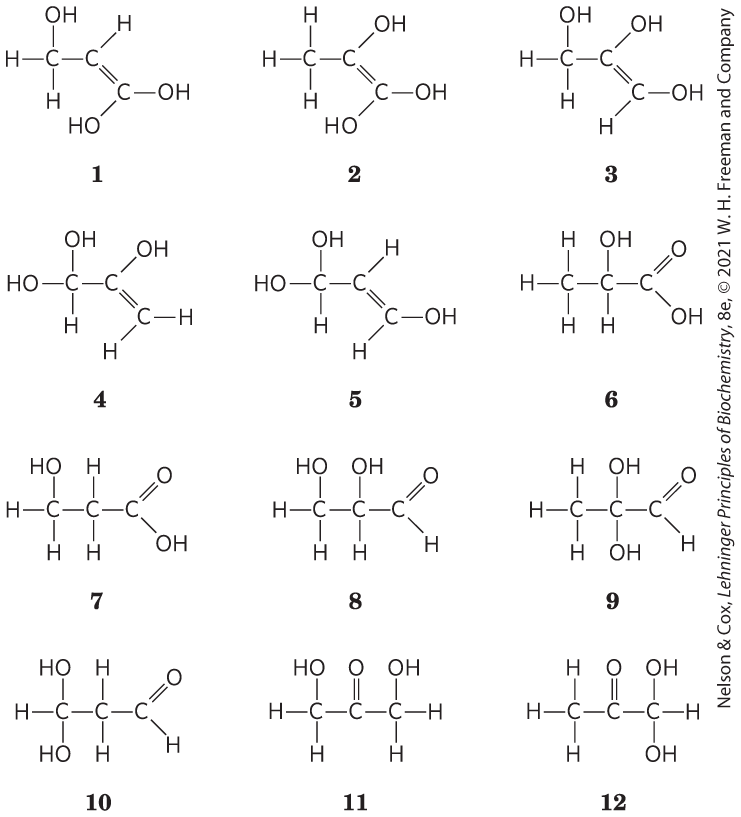
X contains a chiral center; eliminates all but 6 and 8.
X contains an acidic functional group; eliminates 8; structure 6 is consistent with all data.
Structure 6; we cannot distinguish between the two possible enantiomers.
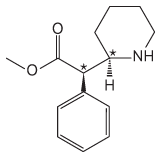
14.
The carbon bearing the hydroxyl group is the chiral carbon.
The structure shows the (R) isomer of propranolol.
(S)-Propranolol has this structure:
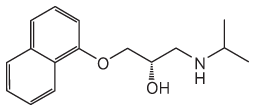
15.
The chiral carbons are indicated with asterisks.
The structure shows the (S,S) isomer of methylphenidate.
(R,R)-methylphenidate has this structure:
16. Spores are alive because they can transition from a state of being metabolically inert under harsh environmental conditions to an actively growing state when conditions improve. During this transition, spores take in water, which is essential to many biochemical reactions. Germination does not seem to depend on ATP.
17.
is negative and relatively large.
There is not enough thermal energy in the wood to overcome the activation energy.
A match supplies thermal energy to overcome the activation energy barrier.
The enzyme lowers the activation energy enough for the reaction to occur at room temperature.
18. If this mutation occurs in a coding sequence, it may cause an amino acid substitution during protein synthesis. The consequence to the cell could be anywhere on the spectrum from beneficial, to neutral, to fatal.
19. The resulting protein may fold incorrectly and may not attain its native conformation. Alternatively, the shape of the region that binds with its partner may change, preventing complementary fit. A mutation that malforms an enzyme’s active site can destroy the enzyme’s catalytic activity.
20.
The copy begins with already-formed domains with biological activity, so it does not have to evolve from scratch. The copy can undergo mutation without harm to the cell, since the original gene encodes the original product. Alternatively, the copy could undergo mutations that create a lethal product, even though the original gene remains intact. The daughter cell’s newly acquired function may be positive or negative.
Each time a duplicate gene undergoes mutation is an opportunity for the cell to acquire a new, positive trait without risking the loss of function provided by the original gene, which may remain unchanged.
21. Yes, when environmental conditions improve, such as temperature moderation and the availability of water, the animal can rehydrate and recover. That is, biochemical mechanisms exist for restoring the normal state.
22. Mutations may have made DNA repair more efficient. Resistant cells may have developed or increased the ability to synthesize a compound that destroys free radicals. Gene duplication may have provided backup for genes damaged by radiation.
23.
The enzyme that catalyzes the reaction is a membrane-bound enzyme because the preparation containing only membranes had the largest amount of labeled product.
The enzyme generated the largest amount of product at pH 7.
Enzyme activity is slightly higher at pH 8 than at pH 6.
Magnesium is effective at activating the system over a range of concentrations. At low concentrations, manganese is effective at activating the system, but it may inhibit the reaction at higher concentrations. The system shows minimal response to calcium ions .
The reaction requires CTP, but not GTP, UTP, or ATP. However, the addition of ATP may increase the rate of lecithin synthesis. The ATP from lot 116 was likely contaminated with CTP.
Chapter 2
1. Weaker; ionic attractive force is proportional to the inverse of the dielectric constant, and a hydrophobic “solvent” such as the environment inside the protein has a lower dielectric constant than a polar solvent such as water.
2. Biomolecular interactions generally need to be reversible; weak interactions allow reversibility.
3. Ethanol is polar; ethane is not. The ethanol group can hydrogen-bond with water.
4.
4.76
9.19
4.0
4.82
5.
6. 1.1
7.
3.3
9.8
8. 1.1
9. 1.7 nmol of acetylcholine
10. 0.1 m hydrofluoric acid
11.
strong
weak
strong
strong
weak
weak
12. 3.3 mL
13.
14.
5.06
4.28
5.46
4.76
3.76
15. quinoline ion: 0.1 m HCl; m-cresol: 0.1 m NaOH; 2-(methylthio)pyridine ion: 0.1 m HCl
16.
Bicarbonate, a weak base, titrates to , making the compound more polar and more water-soluble.
17. Stomach; the neutral form of aspirin present at the lower pH is less polar and passes through the membrane more easily.
18. 8.8
19. 7.4
20.
pH 8.6 to 10.6
4/5
10 mL
21. 8.9
22. 2.4
23. 6.9
24. 1.4
25.
26.
27. Mix 150 mL of 0.10 m sodium acetate and 850 mL of 0.10 m acetic acid.
28.
pH 3
pH 5
pH 9
pH 9
pH 3
pH 5. The total buffering region spans approximately 1 pH unit on either side of the value.
29.
4.6
0.1 pH unit
4 pH units
30. 4.3
31. 0.13 m acetate and 0.07 m acetic acid
32. 1.8
33. 7
34.
fully protonated
zwitterionic
zwitterionic
zwitterionic
fully deprotonated. When the pH is lower than both values, both the -amino group and the -carboxyl group are protonated. When the pH is greater than both values, neither the -amino group nor the -carboxyl group is protonated. When the pH is between the two values, the -amino group is protonated, whereas the -carboxyl group is unprotonated.
35.
Blood pH is controlled by the carbon dioxide–bicarbonate buffer system, . During hypoventilation, increases in the air space of the lungs and arterial blood, driving the equilibrium to the right, raising and lowering blood pH.
During hyperventilation, decreases in the lungs and arterial blood, reducing and increasing pH above the normal 7.4 value.
Lactate is a moderately strong acid, completely dissociating under physiological conditions and thus lowering the pH of blood and muscle tissue. Hyperventilation removes , raising the pH of blood and tissues in anticipation of the acid buildup.
36. 7.4
37. Dissolving more in the blood increases in blood and extracellular fluids, lowering
38.
number of hydrogen bonds that the compound can form and length of the carbon chain
39. The average bond duration decreases.
40.
forms hydrogen bonds with but not with itself.
has a lower boiling point than .
No, is a less polar solvent than .
41. sodium acetate > sodium propionate > glycine > l-phenylalanine > sodium octanoate
42.
43.
44.
No, because an effluent of pH 1 would harm the trout and other life in the stream.
The pH scale runs from very acidic to very alkaline, with the point of neutrality (which is best for living creatures, including trout) midway between 0 and 14. He is proposing to jump out of the frying pan into the fire!
45.
46.
-
47.
Use the substance in its surfactant form to emulsify the spilled oil, collect the emulsified oil, then switch to the nonsurfactant form. The oil and water will separate, and the oil can be collected for further use.
The equilibrium lies strongly to the right. The stronger acid (lower ), , donates a proton to the conjugate base of the weaker acid (higher ), amidine.
The strength of a surfactant depends on the hydrophilicity of its head groups: the more hydrophilic, the more powerful the surfactant. The amidinium form of s-surf is much more hydrophilic than the amidine form, so it is a more powerful surfactant.
Point A: amidinium; the has had plenty of time to react with the amidine to produce the amidinium form. Point B: amidine; Ar has removed from the solution, leaving the amidine form.
The conductivity rises as uncharged amidine reacts with to produce the charged amidinium form.
The conductivity falls as Ar removes , shifting the equilibrium to the uncharged amidine form.
Treat s-surf with to produce the surfactant amidinium form, and use this to emulsify the spill. Treat the emulsion with Ar to remove the and produce the nonsurfactant amidine form. The oil will separate from the water and can be recovered.
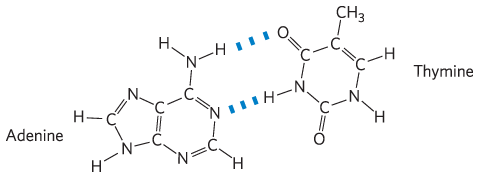
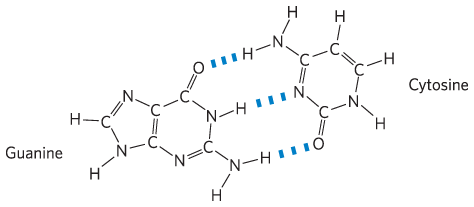
Chapter 3
1. The constituents are Glu, Cys, and Gly. The Glu links to the Cys via its group.
2. It is , because the amino group occupies the same relative position as the hydroxyl group in .
3.
II
IV
I
III
II
II
IV
III
V
III
II and IV
4.
of the -carboxyl group and of the -amino group, so both groups are charged (ionized).
1 in . The pI of alanine is 6.01. From Table 3-1 and the Henderson-Hasselbalch equation, 1 in 4,680 carboxyl groups and 1 in 4,680 amino groups are uncharged. The fraction of alanine molecules with both groups uncharged is 1 in .
5.
b., c.
pH Structure identified in (a) Net charge Migrates toward 1 1 Cathode 4 2 Cathode 8 3 0 Does not migrate 12 4 Anode
6.
Glutamate
Methionine
Aspartate
Glycine
Serine
7.
2
4
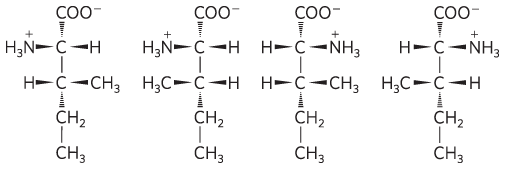
8.
Structure at pH 7:
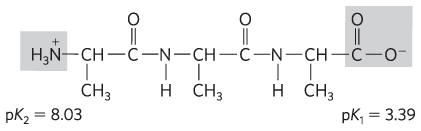
Electrostatic interaction between the carboxylate anion and the protonated amino group of the alanine zwitterion favorably affects ionization of the carboxyl group. This favorable electrostatic interaction decreases as the length of the poly(Ala) increases, resulting in an increase in .
Ionization of the protonated amino group destroys the favorable electrostatic interaction noted in (b). With increasing distance between the charged groups, removal of the proton from the amino group in poly(Ala) becomes easier and thus is lower. The intramolecular effects of the amide (peptide bond) linkages keep values lower than they would be for an alkyl-substituted amine.
9. One H comes from the -amino group of one amino acid, and an OH is removed from the -carboxyl group of the amino acid to which the first is joined.
10. 75,000
11.
10,300. The elements of water are lost when a peptide bond forms, so the molecular weight of a Cys residue is not the same as the molecular weight of free cysteine.
21
12. The protein has four subunits, with molecular masses of 160, 90, 90, and 60 kDa. The two 90 kDa subunits (possibly identical) are linked by one or more disulfide bonds.
13.
at pH 3, ; at pH 8, 0; at pH 11,
14. Lys, His, Arg; negatively charged phosphate groups in DNA interact with positively charged side groups in histones.
15.
16.
Specific activity after step 1 is 6.8 units/mg; step 2, 13 units/mg; step 3, 14 units/mg; step 4, 700 units/mg; step 5, 3,500 units/mg; step 6, 5,000 units/mg.
Step 4
Step 3
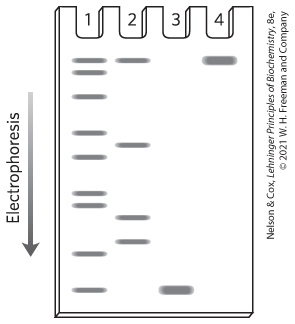
Yes. Specific activity increased only modestly in step 6; SDS polyacrylamide gel electrophoresis.
17.
18. B elutes first, A second, C last.
19. The chymotrypsin protein has three distinct polypeptide chains linked by disulfide bonds. They move on the gel as separate species once the disulfide bonds are broken to form the three peptides in lane 2.
20.
Amino terminus
Tyr–Gly–Gly–Phe–Leu
21. Phosphorylation of serine would alter the mass by 80.
22.
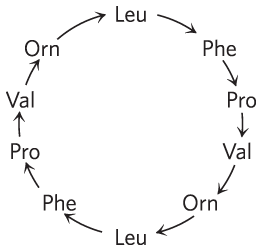
The arrows correspond to the orientation of the peptide bonds, .
23. 75%, 93%. If the efficiency of each amino acid addition is x, then the percentage of full-length peptides with the correct sequence after the addition of seven amino acids will be , as there are seven peptide bonds.
24.
Y (Tyr) at position 1, F (Phe) at position 7, and R (Arg) at position 9.
Positions 4 and 9; K (Lys) is more common at 4, R (Arg) is invariant at 9.
Positions 5 and 10; E (Glu) is more common at both positions.
Position 2; S (Ser).
25.
Anion-exchange chromatography: peptide 2; cation-exchange chromatography: peptide 1; size-exclusion chromatography: peptide 2
peptide 3
26.
Any linear polypeptide chain has only two kinds of free amino groups: a single -amino group at the amino terminus, and an -amino group on each Lys residue. These amino groups react with FDNB to form a DNP–amino acid derivative. Insulin gave two different -amino-DNP derivatives, suggesting that it has two amino termini and thus two polypeptide chains — one with an amino-terminal Gly and the other with an amino-terminal Phe. Because the DNP-lysine product is -DNP-lysine, the Lys is not at an amino terminus.
Yes. The A chain has amino-terminal Gly, the B chain has amino-terminal Phe, and (nonterminal) residue 29 in the B chain is Lys.
Phe–Val–Asp–Glu–. Peptide B1 shows that the amino-terminal residue is Phe. Peptide B2 also includes Val, but since no DNP-Val is formed, Val is not at the amino terminus; it must be on the carboxyl side of Phe. Thus the sequence of B2 is DNP-Phe–Val. Similarly, the sequence of B3 must be DNP-Phe–Val–Asp, and the sequence of the A chain must begin Phe–Val–Asp–Glu–.
No. The known amino-terminal sequence of the A chain is Phe–Val–Asn–Gln–. The Asn and Gln appear in Sanger’s analysis as Asp and Glu because the vigorous hydrolysis in step 7 hydrolyzed the amide bonds in Asn and Gln (as well as the peptide bonds), forming Asp and Glu. Sanger et al. could not distinguish Asp from Asn or Glu from Gln at this stage in their analysis.
The sequence exactly matches that in Fig. 3-24. Each peptide in the table gives specific information about which Asx residues are Asn or Asp and which Glx residues are Glu or Gln.
Ac1: residues 20–21. This is the only Cys–Asx sequence in the A chain; there is amido group in this peptide, so it must be Cys–Asn:
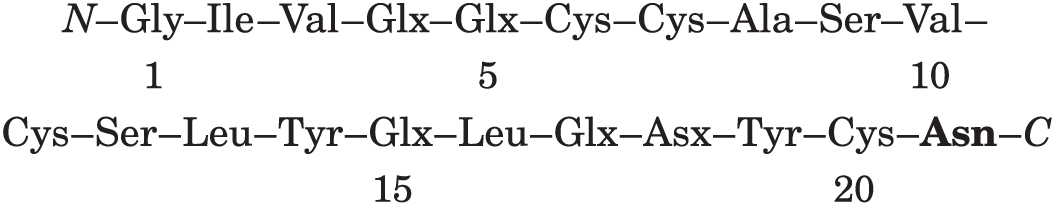
Ap15: residues 14–15–16. This is the only Tyr–Glx–Leu sequence in the A chain; there is amido group, so the peptide must be Tyr–Gln–Leu:
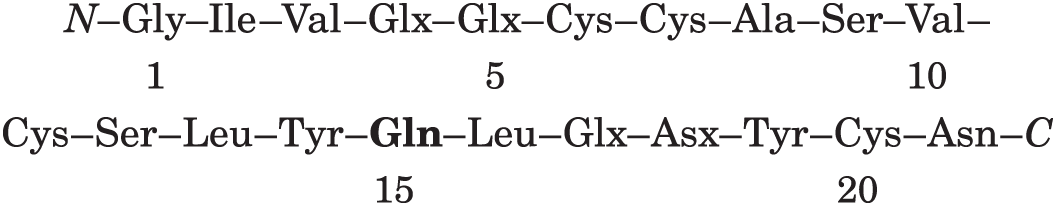
Ap14: residues 14–15–16–17. There is amido group, and we already know that residue 15 is Gln, so residue 17 must be Glu:
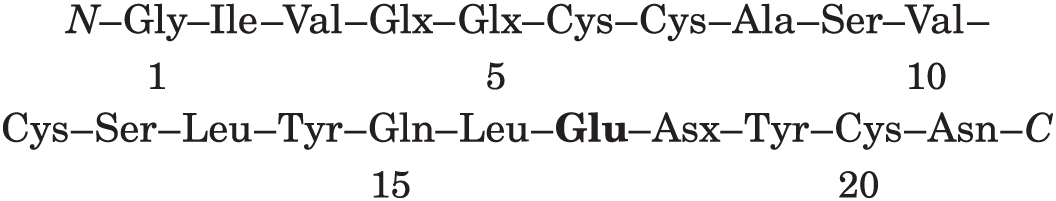
Ap3: residues 18–19–20–21. There are amido groups, and we know that residue 21 is Asn, so residue 18 must be Asn:
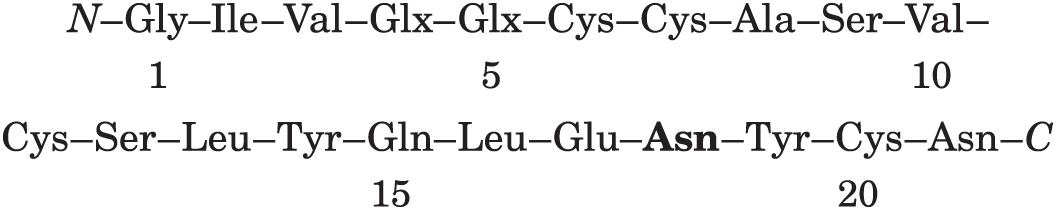
Ap1: residues 17–18–19–20–21, which is consistent with residues 18 and 21 being Asn.
Ap5pa1: residues 1–2–3–4. There are amido groups, so residue 4 must be Glu:
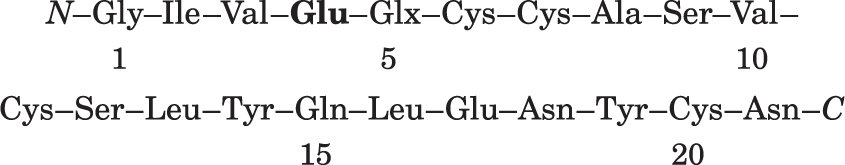
Ap5: residues 1 through 13. There is amido group, and we know that residue 4 is Glu, so residue 5 must be Gln:
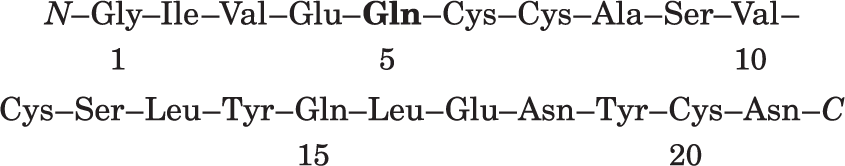
Chapter 4
1.
Shorter bonds have a higher bond order (are multiple rather than single) and are stronger. The peptide C―N bond is stronger than a single bond and is midway between a single bond and a double bond in character.
Rotation about the peptide bond is difficult at physiological temperatures because of its partial double-bond character.
2.
The principal structural units in the wool fiber polypeptide (α-keratin) are successive turns of the helix, at 5.4 Å intervals; coiled coils produce the 5.2 Å spacing. Steaming and stretching the fiber yields an extended polypeptide chain with the conformation, with a distance between adjacent R groups of about 7.0 Å. As the polypeptide reassumes an -helical structure, the fiber shortens.
Wool shrinks in the presence of moist heat, as polypeptide chains are converted from an extended conformation to the native -helix conformation. The structure of silk— sheets, with their small, closely packed amino acid side chains—is more stable than that of wool.
3. peptide bonds per second
4. At the carboxyl groups of poly(Glu) are deprotonated; repulsion among negatively charged carboxylate groups leads to unfolding. Similarly, at pH 7, the amino groups of poly(Lys) are protonated; repulsion among these positively charged groups also leads to unfolding.
5.
Disulfide bonds are covalent bonds, which are much stronger than the noncovalent interactions that stabilize most proteins. They cross-link protein chains, increasing their stiffness, mechanical strength, and hardness.
Cystine residues (disulfide bonds) prevent the complete unfolding of the protein.
6. and
7.
Bends are most likely at residues 7 and 19; Pro residues in the cis configuration accommodate turns well.
The Cys residues at positions 13 and 24 can form disulfide bonds.
External surface: polar and charged residues (Asp, Gln, Lys); interior: nonpolar and aliphatic residues (Ala, Ile); Thr, though polar, has a hydropathy index near zero and thus can be found either on the external surface or in the interior of the protein.
8.
At pH 6.0, the amino acid residues are in the correct protonation state to form an ion pair. At pH 2.0, both Asp and His are predominantly protonated, and at pH 10.0, they are both predominantly deprotonated.
Burial of a charged amino acid residue will destabilize the protein and shift the thermal denaturation curve to lower temperatures.
Lesser. At pH 10.0, a greater fraction of the Lys side chain will be deprotonated and uncharged, facilitating its burial in a hydrophobic environment.
9. 30 amino acid residues; 0.87
10. For many proteins, the amino acid sequence dictates the formation of a unique, folded structure. However, the reverse is not true. Many different amino acid sequences can give rise to similar folded structures. For example, the relative orientation of the charged amino acid residues in an ion pair can be switched while still preserving the overall location of the interaction.
11. Protein (a), a barrel, is described by Ramachandran plot (c), which shows most of the allowable conformations in the upper left quadrant where the bond angles characteristic of the conformation are concentrated. Protein (b), a series of helices, is described by plot (d), where most of the allowable conformations are in the lower left quadrant.
12.
The number of moles of DNP-valine formed per mole of protein equals the number of amino termini and thus the number of polypeptide chains.
4
Different chains would probably run as discrete bands on an SDS polyacrylamide gel.
13.
Aromatic residues seem to play an important role in stabilizing amyloid fibrils. Thus, molecules with aromatic substituents may inhibit amyloid formation by interfering with the stacking or association of the aromatic side chains.
Amyloid forms in the pancreas in association with type 2 diabetes, and forms in the brain in Alzheimer disease. Although the amyloid fibrils in the two diseases involve different proteins, the fundamental structure of the amyloid is similar and is similarly stabilized in both, so they are potential targets for similar drugs designed to disrupt this structure.
14. Although a protein may have only one unique folded structure, many different unfolded structures may exist. The different folding pathways and structures used by the product of other disease alleles of CFTR may not be corrected by lumacaftor.
15.
x-ray crystallography
cryo-EM
NMR
NMR or CD
16.
2QYC is B; 2BNH is C; 2Q5R is E or M; 1XU9 is H; 3H7X is I; 1OU5 is E or M; 2WCD is O.
dimer, monomer, dimer, tetramer, trimer, all dimer, 24-mer or double dodecamer (12-mer), all .
BIOCHEM
17.
NF transcription factor, also called RelA transforming factor
No. You will obtain similar results, but with additional related proteins listed.
The protein has two subunits. There are multiple variants of the subunits, with the best characterized being 50, 52, or 65 kDa. These pair with each other to form a variety of homodimers and heterodimers. The structures of a number of different variants can be found in the PDB.
The NFB transcription factor is a dimeric protein that binds specific DNA sequences, enhancing transcription of nearby genes. One such gene is the immunoglobulin (kappa) light chain, from which the transcription factor gets its name.
18.
Aba is a suitable replacement because Aba and Cys have side chains that are approximately the same size and are similarly hydrophobic. However, Aba cannot form disulfide bonds, so it will not be a suitable replacement if these are required.
There are many important differences between the synthesized protein and HIV protease produced by a human cell, any of which could result in an inactive synthetic enzyme. (1) Although Aba and Cys have a similar size and hydrophobicity, Aba may not be similar enough for the protein to fold properly. (2) HIV protease may require disulfide bonds for proper functioning. (3) Many proteins synthesized by ribosomes fold while being produced; the protein in this study folded only after the chain was complete. (4) Proteins synthesized by ribosomes may interact with the ribosomes as they fold; this is not possible for the protein in the study. (5) Cytosol is a more complex solution than the buffer used in the study; some proteins may require specific, unknown proteins for proper folding. (6) Proteins synthesized in cells often require chaperones for proper folding; these are not present in the study buffer. (7) In cells, HIV protease is synthesized as part of a larger chain that is then proteolytically processed; the protein in the study was synthesized as a single molecule.
Because the enzyme is functional when Aba is substituted for Cys, disulfide bonds do not play an important role in the structure of HIV protease.
Model 1: It would fold like the l-protease. For: The covalent structure is the same (except for chirality), so it should fold like the l-protease. Against: Chirality is not a trivial detail; three-dimensional shape is a key feature of biological molecules. The synthetic enzyme will not fold like the l-protease. Model 2: It would fold to the mirror image of the l-protease. For: Because the individual components are mirror images of those in the biological protein, it will fold in the mirror-image shape. Against: The interactions involved in protein folding are very complex, so the synthetic protein will most likely fold in another form. Model 3: It would fold to something else. For: The interactions involved in protein folding are very complex, so the synthetic protein will most likely fold in another form. Against: Because the individual components are mirror images of those in the biological protein, it will fold in the mirror-image shape.
Model 2. The enzyme is active, but with the enantiomeric form of the biological substrate, and it is inhibited by the enantiomeric form of the biological inhibitor. This is consistent with the d-protease being the mirror image of the l-protease.
Evans blue is achiral; it binds to both forms of the enzyme.
No. Because proteases contain only l-amino acids and recognize only l-peptides, chymotrypsin would not digest the d-protease.
Not necessarily. Depending on the individual enzyme, any of the problems listed in (b) could result in an inactive enzyme.
Chapter 5
1. Protein B has a higher affinity for ligand X; its lower indicates that protein B will be half-saturated at a much lower concentration of X than will protein A. Protein A has ; protein B has .
2.
can mean that the protein exhibits negative cooperativity, where the binding of one ligand to the protein decreases its affinity for other ligand molecules.
b., c. There are some instances where occurs without true negative cooperativity. For example, could also occur if a single polypeptide contained multiple binding sites that had a different affinity for the ligand. If a protein preparation contains a heterogeneous mixture of the protein where some molecules are partially denatured, the measured binding affinity would be artificially decreased, resulting in .
If a protein has multiple ligand binding sites that all have the same binding affinity and do not affect one another, no cooperativity, positive or negative, will be observed.
3.
4.
33 nm
0.15
1.9
5.
(shortcut: the is equivalent to the ligand concentration where ).
The rat receptor has the highest affinity, as it has the lowest .
6. Tight binding of CO to a few binding sites in a hemoglobin tetramer tends to force the entire protein into the R state. can still bind to the unoccupied sites, but it will bind tightly and not be released into the tissues.
7. The cooperative behavior of hemoglobin arises from subunit interactions.
8.
Shift the curve to the right.
Shift the curve to the right.
Shift the curve to the right. All of these conditions would decrease the affinity of hemoglobin for .
9.
The observation that HbA (maternal) is about 60% saturated when the is 4 kPa, whereas HbF (fetal) is more than 90% saturated under the same physiological conditions, indicates that HbF has a higher affinity than HbA.
The higher affinity of HbF ensures that oxygen will flow from maternal blood to fetal blood in the placenta. Fetal blood approaches full saturation where the affinity of HbA is low.
The observation that the -saturation curve of HbA undergoes a larger shift on BPG binding than that of HbF suggests that HbA binds BPG more tightly than does HbF. Differential binding of BPG to the two hemoglobins may determine the difference in their affinities.
10.
Hb Memphis
HbS, Hb Milwaukee, Hb Providence, and possibly Hb Cowtown
Hb Providence
11. More tightly. An inability to form tetramers would limit the cooperativity of these variants, and the binding curve would become more hyperbolic. Also, the BPG-binding site would be disrupted. Oxygen binding would probably be tighter, because the default state in the absence of bound BPG is the tight-binding R state.
12.
. Note that a rearrangement of Eqn 5-8 gives .
13. The epitope is likely to be a structure that is buried when G-actin polymerizes to form F-actin.
14.
The human immune system requires several days to mount a staged response to antigens on the surface of a pathogen. Both trypanosomes and HIV evade the immune system by altering the surface proteins to which immune system components initially bind. Thus, the host organism regularly faces new antigens and requires time to mount an immune response to each one, giving the pathogen time to replicate and spread. HIV also evades the immune system by actively infecting and destroying immune system cells—namely, helper T cells ( cells).
15. Binding of ATP to myosin triggers dissociation of myosin from the actin thin filament. In the absence of ATP, actin and myosin bind tightly to each other.
16.
2
3
1
4
17.
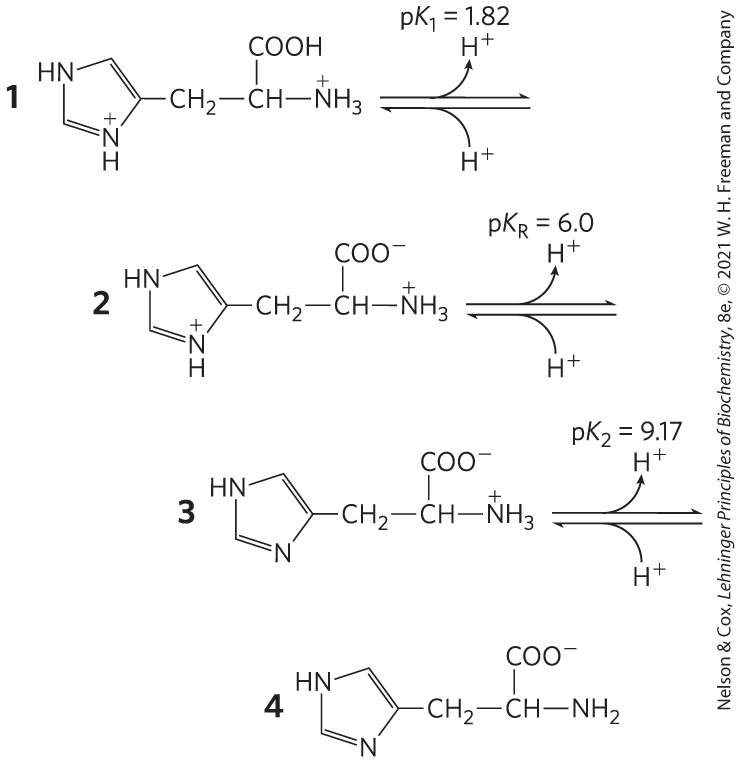
Chain L is the light chain and chain H is the heavy chain of the Fab fragment. Chain Y is lysozyme.
conformation structures are predominant in the variable and constant regions of the fragment.
Fab heavy-chain fragment: 218 amino acid residues; light-chain fragment: 214; lysozyme: 129. Less than 15% of the lysozyme molecule is in contact with the Fab fragment.
Residues that seem to be in contact with lysozyme include, in the H chain: , , , , and ; in the L chain: , , , and . In lysozyme, residues , , , , , , , , , and seem to be situated at the antigen-antibody interface. Not all of these residues are adjacent in the primary structure. Folding of the polypeptide into higher levels of structure brings nonconsecutive residues together to form the antigen-binding site.
18.
2
Instantly. Suitable antibodies are almost always present before any challenge from the virus.
>100,000,000
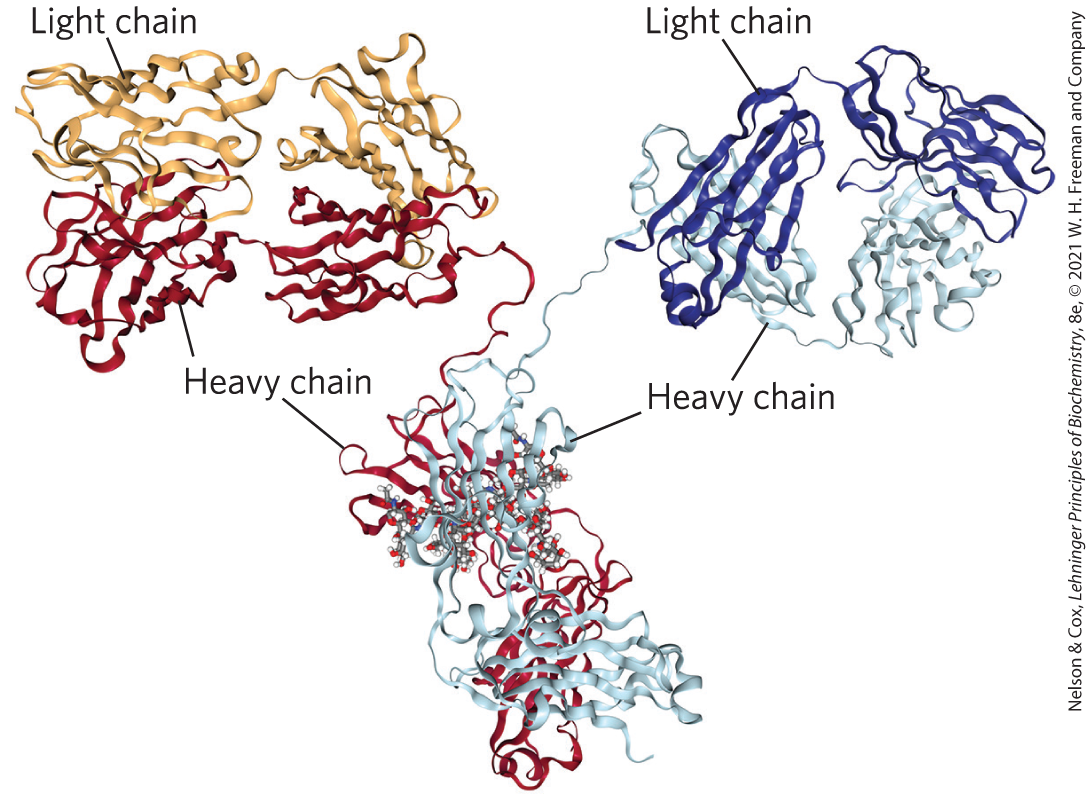
19.
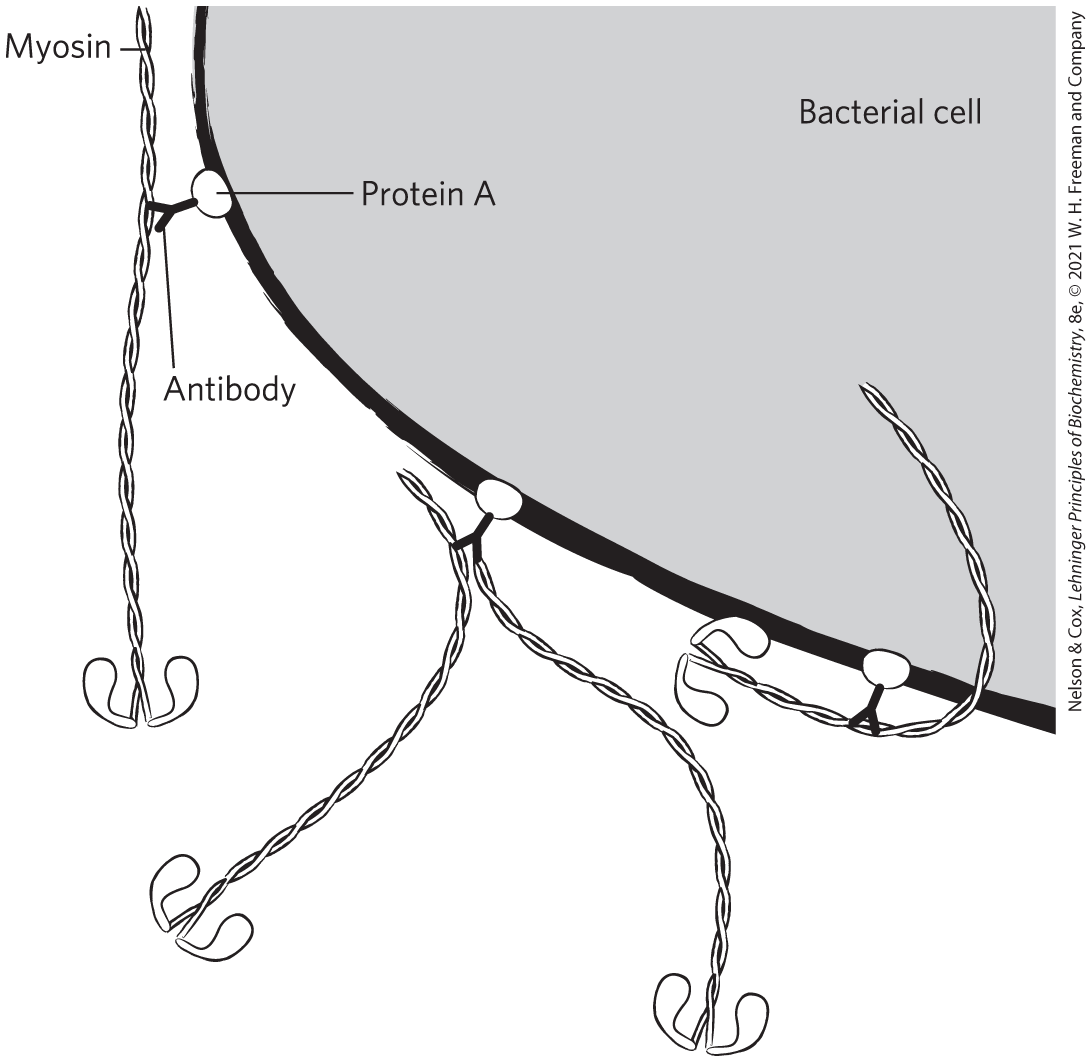
The drawing is not to scale; any given cell would have many more myosin molecules on its surface.
ATP is needed to provide the chemical energy to drive the motion (see Chapter 13).
An antibody that bound to the myosin tail, the actin-binding site, would block actin binding and prevent movement. An antibody that bound to actin would also prevent actin-myosin interaction and thus movement.
There are two possible explanations: (1) Trypsin cleaves only at Lys and Arg residues (see Table 3-6), so would not cleave at many sites in the protein. (2) Not all Arg or Lys residues are equally accessible to trypsin; the most-exposed sites would be cleaved first.
The S1 model. The hinge model predicts that bead-antibody-HMM complexes (with the hinge) would move, but bead-antibody-SHMM complexes (no hinge) would not. The S1 model predicts that because both complexes include S1, both would move. The finding that the beads move with SHMM (no hinge) is consistent only with the S1 model.
With fewer myosin molecules bound, the beads could temporarily fall off the actin as a myosin let go of it. The beads would then move more slowly, as time is required for a second myosin to bind. At higher myosin density, as one myosin lets go, another quickly binds, leading to faster motion.
Above a certain density, what limits the rate of movement is the intrinsic speed with which myosin molecules move the beads. The myosin molecules are moving at a maximum rate; adding more will not increase speed.
Chapter 6
1. The activity of the enzyme that converts sugar to starch is destroyed by heat denaturation.
2.
3. years
4. The enzyme-substrate complex is more stable than the enzyme alone.
5. The reaction rate can be measured by following the decrease in absorption by NADH (at 340 nm) as the reaction proceeds. Determine the value; using substrate concentrations well above the measure initial rate (rate of NADH disappearance with time, measured spectrophotometrically) at several known enzyme concentrations, and plot initial rate versus concentration of enzyme. The plot should be linear, with a slope that provides a measure of LDH concentration.
6. (a), (b), (e)
7.
0.33, 0.67, 0.91
The red curve corresponds to enzyme B ( for this enzyme); the black curve, to enzyme A.
8.
9.
Measured ;
10.
Mixed inhibitor
11.
24 nm
( is exactly one-half so )
( is exactly one-half so times in the presence of inhibitor)
No. well below the diffusion-controlled limit.
12. ;
13.
;
Competitive inhibition
14. ;
15.
A
B
16.
17.
P
Q
E
B
A
F
18.
Increase
Decrease
No change
No change
19. The basic assumptions of the Michaelis-Menten equation still hold. The reaction is at steady state, and the rate is determined by . The equations needed to solve for [ES] are
[E] can be obtained by rearranging Eqn 6-19. The rest follows the pattern of the Michaelis-Menten equation derivation in the text.
20. 29,000. The calculation assumes that there is only one essential Cys residue per enzyme molecule.
21. Activity of the prostate enzyme equals total phosphatase activity in a blood sample minus phosphatase activity in the presence of enough tartrate to completely inhibit the prostate enzyme.
22. The inhibition is mixed. Because seems not to change appreciably, this could be the special case of mixed inhibition called noncompetitive.
23. Apparent . The [S] at which is obtained when all terms except on the right side of Eqn 6-30—that is, —equal . Begin with and solve for [S].
24. An amino acid with a positively charged side chain, such as Lys, His, or Arg, could pull electron density away from Tyr.
25. At pH 5.2, is protonated and neutral and is unprotonated and negatively charged. The optimum activity occurs when is protonated and is unprotonated. Optimal activity requires that be protonated and be unprotonated. Activity thus declines with decreasing pH as is protonated, and with increasing pH as is unprotonated.
26.
Although it can be ionized, the sulfhydryl group is relatively nonpolar (see Chapter 3). Amino butyric acid has a nonpolar side chain of similar size to the Cys side chain.
Because the peptide is made up of d-amino acids, the same shape is unlikely. A mirror image shape is likely, as the d-amino acids are mirror images of their l-stereoisomers. Prior to testing, a completely inactive form is at least plausible.
The mirror image hypothesis is strongly supported. The substrates and inhibitors made up of d-amino acids are uniquely active.
No. The active site of chymotrypsin is configured to act on peptides made up of l-amino acids.
No. The folding process is complex, and sometimes requires assistance from chaperones and specialized enzymes. Some enzymes and proteins can fold spontaneously into an active protein, but others cannot.
Chapter 7
1. With reduction of the carbonyl oxygen to a hydroxyl group, the chemistry at C-1 and C-3 is the same; the glycerol molecule is not chiral.
2. Epimers differ by the configuration about only one carbon.
d-altrose (C-2), d-glucose (C-3), d-gulose (C-4)
d-idose (C-2), d-galactose (C-3), d-allose (C-4)
d-arabinose (C-2), d-xylose (C-3)
3. To convert α-d-glucose to β-d-glucose, the bond between C-1 and the hydroxyl on C-5 (as in Fig. 7-6) must be broken; to convert d-glucose to d-mannose, either the ―H or ―OH bond on C-2 must be broken. Conversion between chair conformations does not require bond breakage; this is the critical distinction between configuration and conformation.
4.
Both are polymers of d-glucose, but they differ in the glycosidic linkage: for cellulose, for glycogen.
Both are hexoses, but glucose is an aldohexose, fructose a ketohexose.
Both are disaccharides, but maltose has two -linked d-glucose units, and sucrose has -linked d-glucose and d-fructose.
5.
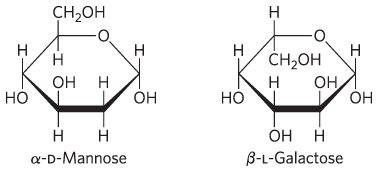
6.
7. An individual with a condition that increases the rate of erythrocyte destruction and turnover would be expected to exhibit less hemoglobin glycation (a lower HbA1c value) because of the shorter-than-normal exposure of the hemoglobin to glucose.
8. A hemiacetal forms when an aldose or a ketose condenses with an alcohol; a glycoside forms when a hemiacetal condenses with an alcohol (see Fig. 7-5).
9. Fructose cyclizes to either the pyranose structure or the furanose structure. Increasing the temperature shifts the equilibrium in the direction of the furanose, the less-sweet form.
10. In 6-phosphogluconolactone, C-1 is a carboxylic acid ester; in glucose, C-1 is a hemiacetal.
11. Boiling a solution of sucrose in water hydrolyzes some of the sucrose to invert sugar. Hydrolysis is accelerated and occurs at lower temperatures with the addition of a small amount of acid (lemon juice or cream of tartar, for example).
12. Prepare a slurry of sucrose and water for the core; add a small amount of sucrase (invertase); immediately coat with chocolate.
13. Sucrose has no free anomeric carbon to undergo mutarotation.
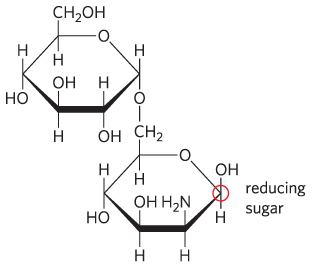
14.
Yes; yes
15. N-Acetyl-β-d-glucosamine is a reducing sugar; its C-1 can be oxidized (p. 237). d-Gluconate is not a reducing sugar; its C-1 is already at the oxidation state of a carboxylic acid. Glc is not a reducing sugar; the anomeric carbons of both monosaccharides are involved in the glycosidic bond.
16. Native cellulose consists of glucose units linked by glycosidic bonds, which force the polymer chain into an extended conformation. Parallel series of these extended chains form intermolecular hydrogen bonds, aggregating into long, tough, insoluble fibers. Glycogen consists of glucose units linked by glycosidic bonds, which cause bends in the chain and prevent formation of long fibers. In addition, glycogen is highly branched and, because many of its hydroxyl groups are exposed to water, is highly hydrated and disperses in water.
Cellulose is a structural material in plants, consistent with its side-by-side aggregation into insoluble fibers. Glycogen is a storage fuel in animals. Highly hydrated glycogen granules with their many nonreducing ends can be rapidly hydrolyzed by glycogen phosphorylase to release glucose 1-phosphate.
17. Cellulose is several times longer; it assumes an extended conformation, whereas amylose has a helical structure.
18. residues/s
19. Glycoproteins: b, c, f; proteoglycans: a, d, e
20. The ball-and-stick models of the disaccharide in Fig. 7-16 show no steric interactions, but space-filling models, showing atoms with their correct relative sizes, would show several strong steric hindrances in the high-energy conformer that are not present in the extended conformer.
21. The negative charges on chondroitin sulfate repel each other and force the molecule into an extended conformation. The polar molecule attracts many water molecules, increasing the molecular volume. In the dehydrated solid, each negative charge is counterbalanced by a positive ion, and the molecule condenses.
22. Positively charged amino acid residues would bind the highly negatively charged groups on heparin. In fact, Lys residues of antithrombin III interact with heparin.
23. The order of the hexoses (ABC, ACB, etc.), the stereochemistry at each of two anomeric carbons (α or β), and the carbon atoms involved in each glycosidic linkage (, etc.)
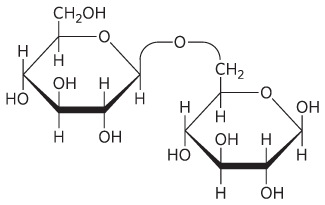
24.
25. Oligosaccharides; their subunits can be combined in more ways than the amino acid subunits of oligopeptides. Each hydroxyl group can participate in glycosidic bonds, and the configuration of each glycosidic bond can be either α or β. The polymer can be linear or branched.
26. Administer an oligosaccharide with the same structure as that recognized by ricin, or high concentrations of N-acetylgalactosamine itself. Ricin will bind the free oligosaccharide or acetylgalactosamine instead of the cell surface target, preventing the entry of the toxin.
27.
Branch-point residues yield 2,3-di-O-methylglucose; unbranched residues yield 2,3,6-tri-O-methylglucose.
3.75%
28.
The tests involve trying to dissolve only part of the sample in a variety of solvents, then analyzing both dissolved and undissolved materials to see whether their compositions differ.
For a pure substance, all molecules are the same and any dissolved fraction will have the same composition as any undissolved fraction. An impure substance is a mixture of more than one compound. When the sample is treated with a particular solvent, more of one component may dissolve, leaving more of the other component(s) behind. As a result, the dissolved and undissolved fractions will have different compositions.
A quantitative assay allows researchers to be sure that none of the activity has been lost through degradation. When the structure of a molecule is being determined, it is important that the sample under analysis consist only of intact (undegraded) molecules. If the sample is contaminated with degraded material, this will give confusing and perhaps uninterpretable structural results. A qualitative assay would detect the presence of activity, even if the sample had become significantly degraded.
Results 1 and 2. Result 1 is consistent with the known structure, because type B antigen has three molecules of galactose; types A and O each have only two. Result 2 is also consistent, because type A has two amino sugars (N-acetylgalactosamine and N-acetylglucosamine); types B and O have only one (N-acetylglucosamine). Result 3 is not consistent with the known structure: for type A, the glucosamine:galactosamine ratio is 1:1; for type B, it is 1:0.
The samples were probably impure and/or partly degraded. The first two results were correct possibly because the method was only roughly quantitative and thus not as sensitive to inaccuracies in measurement. The third result is more quantitative and thus more likely to differ from predicted values because of impure or degraded samples.
An exoglycosidase. If it were an endoglycosidase, one of the products of its action on O antigen would include galactose, N-acetylglucosamine, or N-acetylgalactosamine, and at least one of those sugars would be able to inhibit the degradation. Given that the enzyme is not inhibited by any of these sugars, it must be an exoglycosidase, removing only the terminal sugar from the chain. The terminal sugar of O antigen is fucose, so fucose is the only sugar that could inhibit the degradation of O antigen.
The exoglycosidase removes N-acetylgalactosamine from A antigen and galactose from B antigen. Because fucose is not a product of either reaction, it will not prevent removal of these sugars, and the resulting substances will no longer be active as A antigen or as B antigen. However, the products should be active as O antigen, because degradation stops at fucose.
All the results are consistent with Fig. 10-13. (1) d-Fucose and l-galactose, which would protect against degradation, are not present in any of the antigens. (2) The terminal sugar of A antigen is N-acetylgalactosamine, and this sugar alone protects this antigen from degradation. (3) The terminal sugar of B antigen is galactose, which is the only sugar capable of protecting this antigen.
Chapter 8
1. N-3 and N-7
2. ); it contains a palindrome. The individual strands can form hairpin structures; the two strands can form a cruciform.
3.
4.
Deoxyadenylate, deoxy--methylguanylate, an apurinic site (or AP site or abasic site), deoxyuridylate
end at upper left and end at lower right
The tetranucleotide is DNA, as it is made up of deoxynucleotides. This is true despite the presence of a uracil base.
5. Helices in RNA hairpins assume an A conformation; helices in DNA hairpins generally assume a B conformation.
6. In eukaryotic DNA, about 5% of C residues are methylated. 5-Methylcytosine can spontaneously deaminate to form thymine; the resulting G–T pair is one of the most common mismatches in eukaryotic cells.
7. Higher
8. Without the base, the ribose ring can be opened to generate the noncyclic aldehyde form. This, and the loss of base-stacking interactions, could contribute significant flexibility to the DNA backbone.
9. CGCGCGTGCGCGCGCG
10. RNA nucleotides have a -hydroxyl group on the pentose ring, and the common pyrimidine bases for RNA nucleotides are uracil and cytosine.
11. Base stacking in nucleic acids tends to reduce the absorption of UV light. Denaturation involves loss of base stacking, and UV absorption increases.
12.
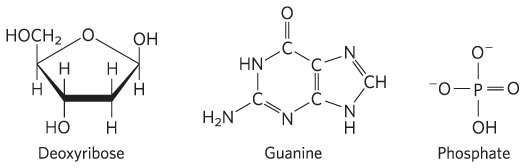
Solubilities: phosphate > deoxyribose > guanine. The highly polar phosphate groups and sugar moieties are on the outside of the double helix, exposed to water; the hydrophobic bases are in the interior of the helix.
13. Primer 1: CCTCGAGTCAATCGATGCTG
Primer 2: CGCGCACATCAGACGAACCA
Recall that all DNA sequences are written in the direction, left to right; that DNA polymerase synthesizes DNA in the direction; that the two strands of a DNA molecule are antiparallel; and that both PCR primers must target the end sequences so that their ends are oriented toward the segment to be amplified.
14.
B
C
A
15. The primers can be used to probe libraries containing long genomic clones to identify contig ends that lie close to each other. If the contigs flanking the gap are close enough, the primers can be used in PCR to directly amplify the intervening DNA separating the contigs, which can then be cloned and sequenced.
16. The -H would prevent addition of any subsequent nucleotides, so the sequence for each cluster would end after the first nucleotide addition.
17. If dCTP is omitted, when the first G residue is encountered in the template, ddCTP will be added, and polymerization will halt. Only one band will be seen in the sequencing gel.
18.
19. The products are:
and the nucleoside -phosphates
20.
Water is a participant in most biological reactions, including those that cause mutations. The low water content in endospores reduces the activity of mutation-causing enzymes and slows the rate of nonenzymatic depurination reactions, which are hydrolysis reactions.
UV light induces formation of cyclobutane pyrimidine dimers. Because B. subtilis is a soil organism, spores can be lofted to the top of the soil or into the air, where they may be subject to prolonged UV exposure.
21. DMT is a blocking group that prevents reaction of the incoming base with itself.
22.
Right-handed. The base at one end is adenine; at the other end, cytosine.
Left-handed
If you cannot see the structures in stereo, use a search engine to find tips online.
23.
It would not be easy! The data for different samples from the same organism show significant variation, and the recovery is never 100%. The numbers for C and T show much more consistency than those for A and G, so for C and T it is much easier to make the case that samples from the same organism have the same composition. But even with the less consistent values for A and G, (1) the range of values for different tissues does overlap substantially; (2) the difference between different preparations of the same tissue is about the same as the difference between samples from different tissues; and (3) in samples for which recovery is high, the numbers are more consistent.
This technique would not be sensitive enough to detect a difference between normal cells and cancerous cells. Cancer is caused by mutations, but these changes in DNA—a few base pairs out of several billion—would be too small to detect with these techniques.
The ratios of A:G and T:C vary widely among different species. For example, in the bacterium Serratia marcescens, both ratios are 0.4, meaning that the DNA contains mostly G and C. In Haemophilus influenzae, by contrast, the ratios are 1.74 and 1.54, meaning that the DNA is mostly A and T.
Conclusion 4 has three requirements. (1) : The table shows an A:T ratio very close to 1 in all cases. Certainly, the variation in this ratio is substantially less than the variation in the A:G and T:C ratios. (2) : Again, the G:C ratio is very close to 1, and the other ratios vary widely. (3) : This is the purine:pyrimidine ratio, which also is very close to 1.
Chapter 9
1.
() – – – G() and ()AATTC – – – ()
() – – – CTTAA() () G – – – ()
() – – – GAATT() and ()AATTC – – – ()
() – – – CTTAA() ()TTAAG – – – ()
() – – – GAATTAATTC – – – ()
() – – – CTTAATTAAG – – – ()
() – – – G() and ()C – – – ()
() – – – C() ()G – – – ()
() – – – GAATTC – – – ()
() – – – CTTAAG – – – ()
() – – – CAG() and ()CTG – – – ()
() – – – GTC() ()GAC – – – ()
() – – – CAGAATTC – – – ()
() – – – GTCTTAAG – – – ()
Method 1: Cut the DNA with EcoRI as in (a), then treat the DNA as in (b) or (d), and then ligate a synthetic DNA fragment with the BamHI recognition sequence between the two resulting blunt ends. Method 2 (more efficient): Synthesize a DNA fragment with the structure
This would ligate efficiently to the sticky ends generated by EcoRI cleavage, would introduce a BamHI site, but would not regenerate the EcoRI site.
The four fragments (with nucleotide), in order of discussion in the problem, are
2. Yeast artificial chromosomes (YACs) are not stable in a cell unless they have two telomere-containing ends and a large DNA segment cloned into the chromosome. YACs less than 10,000 bp long are soon lost during continued mitosis and cell division.
3.
Plasmids in which the original pBR322 was regenerated without insertion of a foreign DNA fragment; these would retain resistance to ampicillin. Also, two or more molecules of pBR322 might be ligated together with or without insertion of foreign DNA.
The clones in lanes 1 and 2 each have one DNA fragment inserted in different orientations. The clone in lane 3 has two DNA fragments, ligated such that the EcoRI proximal ends are joined.
4.
5. Your test would require DNA primers, a heat-stable DNA polymerase, deoxynucleoside triphosphates, and a PCR machine (thermal cycler). The primers would be designed to amplify a DNA segment encompassing the CAG repeat. The DNA strand shown is the coding strand, oriented , left to right. The primer targeted to DNA to the left of the repeat would be identical to any 25-nucleotide sequence shown in the region to the left of the CAG repeat. The primer on the right side must be complementary and antiparallel to a 25-nucleotide sequence to the right of the CAG repeat. Using the primers, DNA including the CAG repeat would be amplified by PCR, and its size would be determined by comparison with size markers after electrophoresis. The length of the DNA would reflect the length of the CAG repeat, providing a simple test for the disease.
6. Design PCR primers that are complementary to the DNA in the deleted segment but would direct DNA synthesis away from each other. No PCR product is generated unless the ends of the deleted segment are joined to create a circle.
7. The two proteins likely co-localize under nutrient starvation and possibly form a protein complex.
8.
9. The production of labeled antibodies is difficult and expensive, and the labeling of every antibody to every protein target would be impractical. By labeling one antibody preparation for binding to all antibodies of a particular class, the same labeled antibody preparation can be used in many different immunofluorescence experiments.
10. Express the protein in yeast strain 1 as a fusion protein with one of the domains of Gal4p—say, the DNA-binding domain. Using yeast strain 2, make a library in which essentially every protein of the fungus is expressed as a fusion protein with the interaction domain of Gal4p. Mate strain 1 with the strain 2 library, and look for colonies that are colored due to expression of the reporter gene. These colonies will generally arise from mated cells containing a fusion protein that interacts with your target protein.
11. Reverse transcriptase is used to convert single-stranded RNA into double-stranded DNA in one of the early steps of RNA-Seq.
12. RNA-Seq detects noncoding RNAs. These have special functions, and they lack protein-coding sequences. Many RNAs encoded by eukaryotic genomes are not messenger RNAs. Instead, they are noncoding RNAs with a variety of functions. They need not possess an open reading frame as part of their sequence.
13. ATSAAGWDEWEGGKVLIHLDGKLQNRGALLELDIGAV
14. The pattern of haplotypes in the Aleut and Eskimo populations suggests that their ancestors’ migration into the American Arctic regions was separate from the migrations that eventually populated the rest of North America and South America.
15. Interbreeding between the Denisovans and Homo sapiens must have occurred in Asia, sometime in the many millennia during which humans migrated from Africa to Asia and then to Australia and Melanesia.
16. The same disease condition can be caused by defects in two or more genes that are on different chromosomes.
17.
would act as a suitable primer pair for this transcript
would form primer dimers because of the high number of complementary bases in the primers
also exhibits significant self-complementarity and would have a very high melting point due to the C–G pairs
also exhibits self-complementarity and would form stem-loops.
18.
DNA solutions are highly viscous because the very long molecules are tangled in solution. Shorter molecules tend to tangle less and form a less viscous solution, so decreased viscosity corresponds to shortening of the polymers—as caused by nuclease activity.
An endonuclease. An exonuclease removes single nucleotides from the end or end and would produce TCA-soluble nucleotides. An endonuclease cuts DNA into oligonucleotide fragments and produces little or no TCA-soluble material.
The end. If the phosphate were left on the end, the kinase would incorporate significant as it added phosphate to the end; treatment with the phosphatase would have no effect on this. In this case, samples A and B would incorporate significant amounts of . When the phosphate is left on the end, the kinase does not incorporate any : it cannot add a phosphate if one is already present. Treatment with the phosphatase removes phosphate, and the kinase then incorporates significant amounts of . Sample A will have little or no , and B will show substantial incorporation—as was observed.
Random breaks would produce a distribution of fragments of random size. The production of specific fragments indicates that the enzyme is site-specific.
Cleavage at the site of recognition. This produces a specific sequence at the end of the fragments. If cleavage occurred near but not within the recognition site, the sequence at the end of the fragments would be random.
The results are consistent with two recognition sequences, as shown below, cleaved where shown by the arrows:
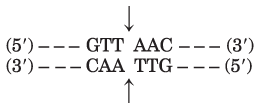
which gives the pApApC and TpTp fragments; and
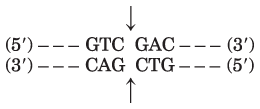
which gives the ()pGpApC and ()CpTp fragments.
Chapter 10
1. The term “lipid” does not specify a particular chemical structure. Compounds are categorized as lipids based on their greater solubility in organic solvents than in water.
2.
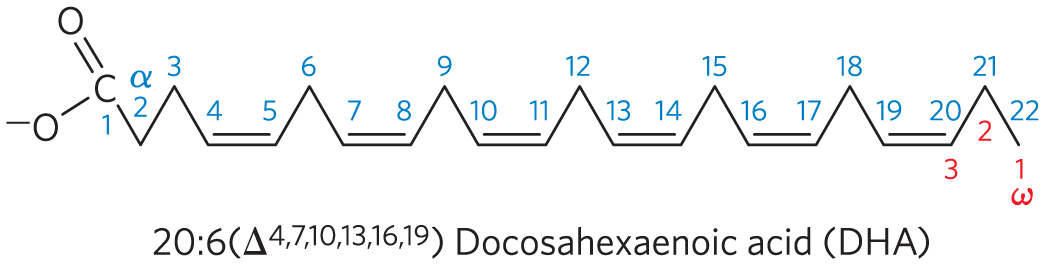
3.
The number of cis double bonds. Each cis double bond causes a bend in the hydrocarbon chain, lowering the melting temperature.
Six different triacylglycerols can be constructed, in order of increasing melting points:
where oleic and palmitic acid. The greater the content of saturated fatty acid, the higher the melting point.
Branched-chain fatty acids increase the fluidity of membranes because they decrease the extent of membrane lipid packing.
4. It reduces double bonds, which increases the melting point of lipids containing the fatty acids.
5. Long, saturated acyl chains, nearly solid at air temperatures, form a hydrophobic layer in which a polar compound such as cannot dissolve or diffuse.
6. Spearmint is (R)-carvone; caraway is (S)-carvone.
7. The label appears in the fatty acid salts.
8. Hydrophobic units:
2 fatty acids
b., c., and d. 1 fatty acid and the hydrocarbon chain of sphingosine
the steroid nucleus and acyl side chain.
Hydrophilic units:
phosphoethanolamine
phosphocholine
d-galactose
several sugar molecules
alcohol group ()
9. Serine
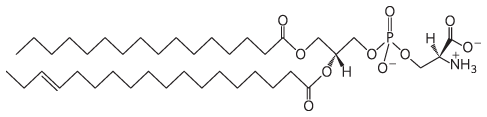
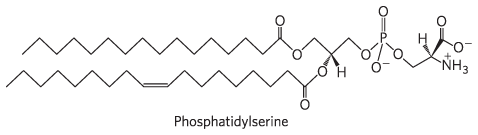
10.
11. The part of the membrane lipid that determines blood type is the oligosaccharide in the head group of the membrane sphingolipids (see Fig. 10-13). This same oligosaccharide is attached to certain membrane glycoproteins, which also serve as points of recognition by the antibodies that distinguish blood groups.
12.
The free group on C-2 and the phosphocholine head group on C-3 are hydrophilic; the fatty acid on C-1 of lysolecithin is hydrophobic.
Certain steroids, such as prednisone, inhibit the action of phospholipase , inhibiting the release of arachidonic acid from C-2. Arachidonic acid is converted to a variety of eicosanoids, some of which cause inflammation and pain.
Phospholipase releases arachidonic acid, a precursor of other eicosanoids with vital protective functions in the body; it also breaks down dietary glycerophospholipids.
13. Diacylglycerol is hydrophobic and remains in the membrane. Inositol 1,4,5-trisphosphate is highly polar, very soluble in water, and more readily diffusible in the cytosol. Both are second messengers.
14.
15.
Glycerol and the sodium salts of palmitic and stearic acids
d-Glycerol 3-phosphocholine and the sodium salts of palmitic and oleic acids
16. Solubility in water: monoacylglycerol > diacylglycerol > triacylglycerol
17. First eluted to last eluted: cholesteryl palmitate and triacylglycerol; cholesterol and n-tetradecanol; phosphatidylcholine and phosphatidylethanolamine; sphingomyelin; phosphatidylserine and palmitate. The lipids elute from the silica gel column in order of polarity. The least polar lipid will elute first and the most polar lipid will elute last.
18.
Subject acid hydrolysates of each compound to chromatography (GC or silica gel TLC), and compare the result with known standards. Sphingomyelin hydrolysate: sphingosine, fatty acids, phosphocholine, choline, and phosphate; cerebroside hydrolysate: sphingosine, fatty acids, sugars, but no phosphate.
Strong alkaline hydrolysis of sphingomyelin yields sphingosine; phosphatidylcholine yields glycerol. Detect hydrolysate components on thin-layer chromatograms by comparing with standards or by their differential reaction with FDNB (only sphingosine reacts to form a colored product). Treatment with phospholipase or releases free fatty acids from phosphatidylcholine, but not from sphingomyelin.
19.
Sphingosine (4.78); linoleic acid (5.88); stearic acid (6.33); cholesterol (7.68)
Log P describes the lipophilicity of the drug, crucial for determining how to formulate the drug for transport through aqueous compartments of the body such as the gut and the bloodstream. Log P also determines the likelihood of a drug being absorbed by fats and fatty tissues, which can alter its effectiveness, half-life, and potential toxicity.
20.
barrel
Phe, Trp, Tyr, Leu. All are hydrophobic or have nonpolar R groups.
The polar head group can hydrogen bond with water; the hydrocarbon tail cannot. Hydrophobic portions of the residues of the pocket protect the tail from contact with water as it moves through the bloodstream.
21.
GM1 and globoside. Both glucose and galactose are hexoses, so “hexose” in the molar ratio refers to glucose + galactose. The ratios for the four gangliosides are GM1, 1:3:1:1; GM2, 1:2:1:1; GM3, 1:2:0:1; globoside, 1:3:1:0.
Yes. The ratio matches GM2, the ganglioside expected to build up in Tay-Sachs disease (see Box 10-1, Fig. 1).
This analysis is similar to that used by Sanger to determine the amino acid sequence of insulin. The analysis of each fragment reveals only its composition, not its sequence, but because each fragment is formed by sequential removal of one sugar, we can draw conclusions about sequence. The structure of the normal asialoganglioside is ceramide–glucose–galactose–galactosamine–galactose, consistent with Box 10-1 (excluding Neu5Ac, removed before hydrolysis).
The Tay-Sachs asialoganglioside is ceramide–glucose–galactose–galactosamine, consistent with Box 10-1.
The structure of the normal asialoganglioside, GM1, is ceramide–glucose [2 involved in glycosidic links; 1 involved in ring structure; 3 (2, 3, 6) free for methylation]–galactose [2 in links; 1 in ring; 3 (2, 4, 6) free for methylation]–galactosamine [2 in links; 1 in ring; 1 instead of ; 2 (4, 6) free for methylation]–galactose [1 in link; 1 in ring; 4 (2, 3, 4, 6) free for methylation].
Two key pieces of information are missing: What are the linkages between the sugars? Where is Neu5Ac attached?
Chapter 11
1. The area per molecule would be calculated from the known amount (number of molecules) of lipid used and the area occupied by a monolayer when it begins to resist compression (when the required force increases dramatically, as shown in the plot of force versus area).
2.
Lipids that form bilayers are amphipathic molecules: they contain a hydrophilic region and a hydrophobic region. To minimize the hydrophobic area exposed to the water surface, these lipids form two-dimensional sheets, with the hydrophilic regions exposed to water and the hydrophobic regions buried in the interior of the sheet. Furthermore, to avoid exposing the hydrophobic edges of the sheet to water, lipid bilayers close on themselves.
These sheets form the closed membrane surfaces that envelop cells and compartments within cells (organelles).
3. 2 nm. Two palmitates placed end to end span about 4 nm, approximately the thickness of a typical bilayer.
4. Integral proteins are firmly embedded in the lipid bilayer and can be released only by treating membranes with a detergent or nonpolar solvent. Peripheral membrane proteins are more easily released, by changes in pH, metal ion concentration, or protein-denaturing reagents like urea. Amphitropic membrane proteins are loosely and reversibly associated with membranes and move between membrane and cytosol as part of their function.
5. Salt extraction indicates a peripheral location, and inaccessibility to protease in intact cells indicates an internal location. Protein X is likely to be a peripheral protein on the cytosolic face of the membrane.
6. Construct a hydropathy plot; hydrophobic regions of 20 or more residues suggest transmembrane segments. Determine whether the protein in intact erythrocytes reacts with a membrane-impermeant reagent specific for primary amines; if it does, the transporter’s amino terminus is on the outside of the cell.
7. ∼4%; estimated by calculating the surface area of the cell and of 10,000 transporter molecules
8. ∼22. To estimate the fraction of membrane surface covered by phospholipids, you would need to know (or estimate) the average cross-sectional area of a phospholipid molecule in a bilayer (e.g., from an experiment such as that described in Problem 1 in this chapter) and the average cross-sectional area of a 50 kDa protein.
9. Rate of diffusion would decrease. Movement of individual lipids in bilayers occurs much faster at 37 ºC, when the lipids are in the “fluid” phase, than at 10 ºC, when they are in the “solid” phase. This effect is more pronounced than the usual decrease in Brownian motion with decreased temperature.
10. Interactions among membrane lipids are due to the hydrophobic effect, noncovalent and reversible, allowing membranes to spontaneously reseal.
11. The temperature of body tissues at the extremities is lower than that of tissues closer to the center of the body. If lipid is to remain fluid at this lower temperature, it must contain a higher proportion of unsaturated fatty acids; unsaturated fatty acids lower the melting point of lipid mixtures.
12. There is a very high energetic barrier to taking the polar head of a membrane lipid through the hydrocarbon core. Higher temperatures might make it more likely to occur, as would the presence of a catalyst, such as a flippase, a floppase, or a scramblase protein.
13. The energetic cost of moving the highly polar, sometimes charged, head group through the hydrophobic interior of the bilayer is prohibitive.
14. Scramblases catalyze the transport of membrane lipids from one membrane leaflet to the other. The reaction is ATP-independent, driven by a transbilayer lipid gradient. Scramblases cannot create an asymmetric distribution of lipids across the bilayer. Flippases catalyze the ATP-dependent transport of aminophospholipids (phosphatidylserine and phosphatidylethanolamine) from the extracellular or lumenal leaflet of a membrane to the cytosolic leaflet.
15. At pH 7, tryptophan bears both a positive charge and a negative charge, but indole is uncharged. The movement of the less polar indole through the hydrophobic core of the bilayer is energetically more favorable.
16. The transporter has a greater than and it is a cotransporter, a symporter with .
17.
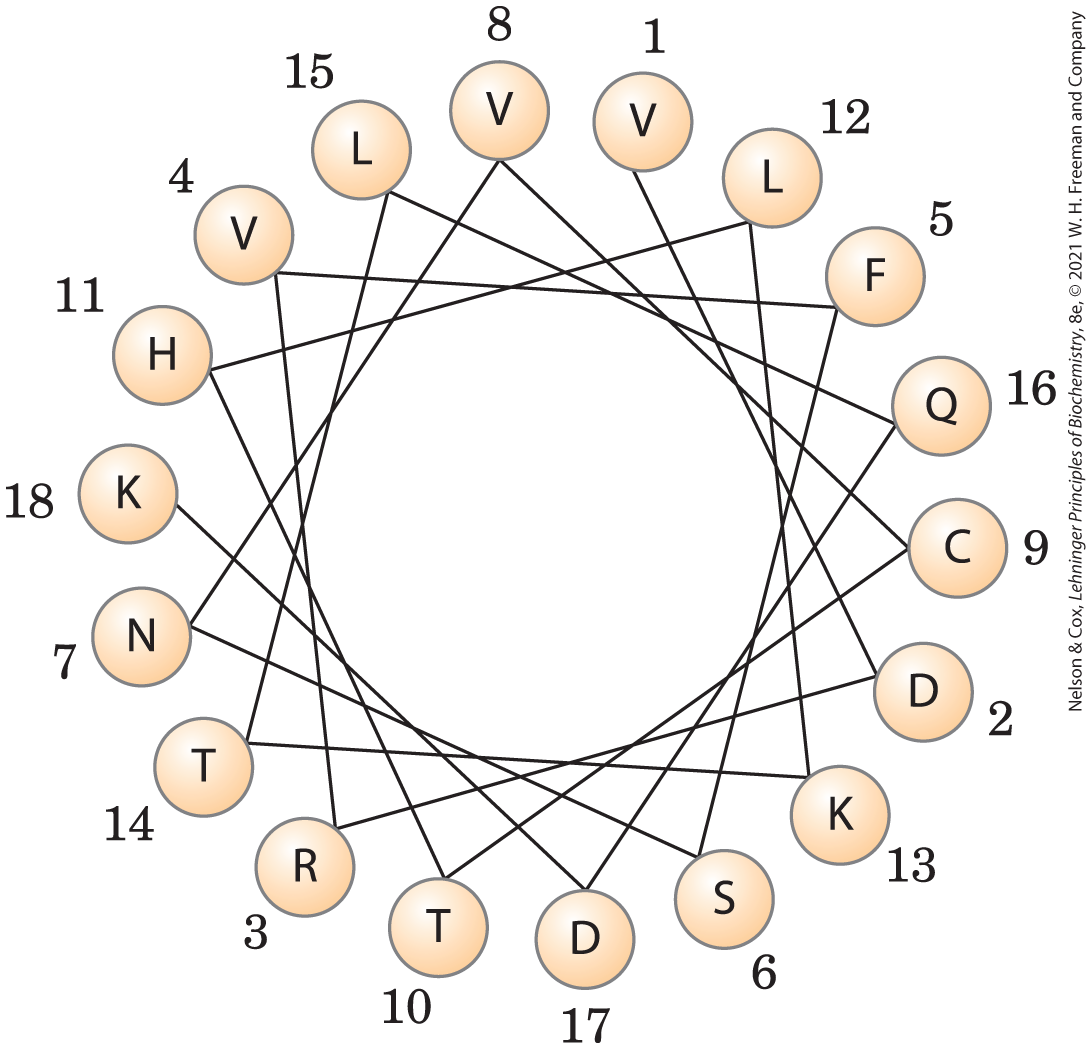
The amino acids with the greatest hydropathy index (V, L, F, and C) are clustered on one side of the helix. This amphipathic helix is likely to dip into the lipid bilayer along its hydrophobic surface while exposing its other surface to the aqueous phase. Alternatively, a group of helices may cluster with their polar surfaces in contact with one another and their hydrophobic surfaces facing the lipid bilayer.
18. 0.60 mol
19. Valinomycin is an ionophore that will carry across the plasma membrane, deflating the membrane potential that is normally achieved by the unequal pumping of and by the ATPase.
20. 13 kJ/mol
21. Most of the consumed by a tissue is for oxidative phosphorylation, the source of most of the ATP. Therefore, about two-thirds of the ATP synthesized by the kidney is used for pumping and .
22. Under normal conditions, the exchanger pumps out as it allows to move inward. The excess of on the outside is created by the ATPase. When that enzyme is blocked with digoxin, the gradient is depleted, and with it, the driving force for exit. So would flow down its gradient, entering the cell. This (increased concentration) is generally lethal for the cell.
23. No. The symporter may carry more than one equivalent of for each mole of glucose transported.
24. Treat a suspension of cells with unlabeled NEM in the presence of excess lactose, remove the lactose, then add radiolabeled NEM. Use SDS-polyacrylamide gel electrophoresis (SDS-PAGE) to determine the of the radiolabeled band (the transporter).
25. ATP-dependence signifies active transport; []-independence suggests primary transport.
26. The leucine transporter is specific for the l isomer, but the binding site can accommodate either l-leucine or l-valine. Reduction of in the absence of indicates that leucine (or valine) is transported by symport with . By depleting the gradient, ouabain would inhibit l-leucine uptake.
27. The channel has a “pore” that allows to diffuse through the channel, stabilized by its interaction with the carbonyl oxygens of the amino acids that line the pore. is smaller than , so it is not sterically hindered from passage through the pore, but is too small to interact with the carbonyl oxygens, so it is not stabilized by that interaction.
28. increases; is unaffected.
29.
Glycophorin A: 1 transmembrane segment; myoglobin: no segments long enough to cross a membrane (not a membrane protein); aquaporin: 6 transmembrane segments (may be a membrane channel or a receptor protein)
The 15 residue window provides a better signal-to-noise ratio.
A narrower window reduces the impact of “edge effects” when a transmembrane sequence occurs near either end of the protein.
30.
The rise per residue for an α helix (Chapter 4) is about . To span a 4 nm bilayer, an α helix must contain about 27 residues; thus, for seven spans, about 190 residues are required. A protein of has about 580 residues.
A hydropathy plot is used to locate transmembrane regions.
Because about half of this portion of the receptor consists of charged residues, it probably represents an intracellular loop that connects two adjacent membrane-spanning regions of the protein.
Because this helix is composed mostly of hydrophobic residues, this portion of the receptor is probably one of the membrane-spanning regions of the protein.
31.
Model A: supported. The two dark lines are either the protein layers or the phospholipid heads, and the clear space is either the bilayer or the hydrophobic core, respectively. Model B: not supported. This model requires a more-or-less uniformly stained band surrounding the cell. Model C: supported, with one reservation. The two dark lines are the phospholipid heads; the clear zone is the tails. This assumes that the membrane proteins are not visible, because they do not stain with osmium or do not happen to be in the sections viewed.
Model A: supported. A “naked” bilayer layers of protein (2 nm) sums to 6.5 nm, which is within the observed range of thickness. Model B: neither. This model makes no predictions about membrane thickness. Model C: unclear. The result is hard to reconcile with this model, which predicts a membrane as thick as, or slightly thicker than (due to the projecting ends of embedded proteins), a “naked” bilayer. The model is supported only if the smallest values for membrane thickness are correct or if a substantial amount of protein projects from the bilayer.
Model A: unclear. The result is hard to reconcile with this model. If the proteins are bound to the membrane by ionic interactions, the model predicts that the proteins contain a high proportion of charged amino acids, in contrast to what was observed. Also, because the protein layer must be very thin (see (b)), there would not be much room for a hydrophobic protein core, so hydrophobic residues would be exposed to the solvent. Model B: supported. The proteins have a mixture of hydrophobic residues (interacting with lipids) and charged residues (interacting with water). Model C: supported. The proteins have a mixture of hydrophobic residues (anchoring in the membrane) and charged residues (interacting with water).
Model A: unclear. The result is hard to reconcile with this model, which predicts a ratio of exactly 2.0; this would be hard to achieve under physiologically relevant pressures. Model B: neither. This model makes no predictions about amount of lipid in the membrane. Model C: supported. Some membrane surface area is taken up with proteins, so the ratio would be less than 2.0, as was observed under more physiologically relevant conditions.
Model A: unclear. The model predicts proteins in extended conformations rather than globular conformations, so is supported only if one assumes that proteins layered on the surfaces include helical segments. Model B: supported. The model predicts mostly globular proteins (containing some helical segments). Model C: supported. The model predicts mostly globular proteins.
Model A: unclear. The phosphorylamine head groups are protected by the protein layer, but only if the proteins completely cover the surface will the phospholipids be completely protected from phospholipase. Model B: supported. Most head groups are accessible to phospholipase. Model C: supported. All head groups are accessible to phospholipase.
Model A: not supported. Proteins are entirely accessible to trypsin digestion, and virtually all will undergo multiple cleavage, with no protected hydrophobic segments. Model B: not supported. Virtually all proteins are in the bilayer and inaccessible to trypsin. Model C: supported. Segments of protein that penetrate or span the bilayer are protected from trypsin; those exposed at the surfaces will be cleaved. The trypsin-resistant portions have a high proportion of hydrophobic residues.
Chapter 12
1. X is cAMP; its production is stimulated by epinephrine.
Centrifugation sediments adenylyl cyclase (which catalyzes cAMP formation) in the particulate fraction.
Added cAMP stimulates glycogen phosphorylase.
cAMP is heat-stable; it can be prepared by treating ATP with barium hydroxide.
2. Unlike cAMP, dibutyryl cAMP passes readily through the plasma membrane.
3.
It increases [cAMP].
cAMP regulates permeability.
Replace lost body fluids and electrolytes.
4.
The mutation makes R unable to bind and inhibit C, so C is constantly active.
The mutation prevents cAMP binding to R, leaving C inhibited by bound R.
5. Albuterol raises [cAMP], leading to relaxation and dilation of the bronchi and bronchioles. Because β-adrenergic receptors control many other processes, this drug would have undesirable side effects. To minimize these effects, find an agonist specific for the subtype of β-adrenergic receptors found in bronchial smooth muscle.
6. Hormone degradation; hydrolysis of GTP bound to a G protein; degradation, metabolism, or sequestration of second messenger; receptor desensitization; removal of receptor from the cell surface.
7. Fuse CFP to β-arrestin and fuse YFP to the cytoplasmic domain of the β-adrenergic receptor, or vice versa. In either case, illuminate at 433 nm and observe fluorescence at both 476 nm and 527 nm. If the interaction occurs, emitted light intensity will decrease at 476 nm and increase at 527 nm upon addition of epinephrine to cells expressing the fusion proteins. If the interaction does not occur, the wavelength of emitted light will remain at 476 nm. Some reasons why this might fail: the fusion proteins (1) are inactive or otherwise unable to interact, (2) are not translocated to their normal subcellular location, or (3) are not stable to proteolytic breakdown.
8. Vasopressin acts by elevating cytosolic to , activating protein kinase C. EGTA injection blocks vasopressin action but should not affect the response to glucagon, which uses cAMP, not , as second messenger.
9. Amplify: (a), (b), (e), (f). Terminate: (c), (d), (f). (f) can contribute to both.
10. IRS1, Grb2, Sos, Ras, Raf, MEK, ERK
11. A mutation in ras that inactivated the Ras GTPase activity would create a protein that, once activated by the binding of GTP, would continue to give, through Raf, the insulin-response signal.
12. Shared properties of Ras and : Both bind either GDP or GTP; both are activated by GTP; both, when active, activate a downstream enzyme; both have intrinsic GTPase activity that shuts them off after a short period of activation. Differences: Ras is a small, monomeric protein; is heterotrimeric. Functional difference between and : activates adenylyl cyclase; inhibits it.
13. Kinase (factor in parentheses): PKA (cAMP); PKG (cGMP); PKC (, DAG); CaM kinase (, CaM); cyclin-dependent kinase (cyclin); receptor Tyr kinase (ligand for the receptor, such as insulin); MAPK (Raf); Raf (Ras); glycogen phosphorylase kinase (PKA).
14. remains in its activated form when the nonhydrolyzable analog is bound. The analog therefore prolongs the effect of epinephrine on the injected cell.
15. Individuals with Oguchi disease might have a defect in rhodopsin kinase or in arrestin.
16. Rod cells would no longer show any change in membrane potential in response to light. This experiment has been done. Illumination did activate PDE, but the enzyme could not significantly reduce the 8-Br-cGMP level, which remained well above that needed to keep the gated ion channels open. Thus, light had no impact on membrane potential.
17. Insulin increases glycogen synthesis.
18. Nearly every component of the β-adrenergic and insulin receptor signaling pathways communicates signal activation by some connection through an IDR. The activation loop of protein kinases is an IDR, and the carboxyl-terminal end of most of the protein kinases in those pathways is an IDR. AKAPs and other scaffold proteins serve as anchors to hold pathway components in proximity. Phosphorylation/dephosphorylation of IDRs serves as a switch for the ability of the target proteins to associate.
19. (b), (c), (e), (d), (a)
20.
On exposure to heat, TRPV1 channels open, causing an influx of and into the sensory neuron. This depolarizes the neuron, triggering an action potential. When the action potential reaches the axon terminus, neurotransmitter is released, signaling to the nervous system that heat has been sensed.
Capsaicin mimics the effects of heat by opening TRPV1 at low temperatures, leading to the false sensation of heat. The extremely low indicates that even very small amounts of capsaicin will have dramatic sensory effects.
At low levels, menthol should open the TRPM8 channel, leading to a sensation of cool; at high levels, both TRPM8 and TRPV3 open, leading to a mixed sensation of cool and heat, such as you may have experienced with very strong peppermints.
21.
These mutations might lead to permanent activation of the receptor, leading to unregulated cell division and tumor formation.
The viral gene might encode a constitutively active form of the receptor, causing a constant signal for cell division and thus tumor formation.
E1A protein might bind to pRb and prevent E2F from binding, so E2F is constantly active and cells divide uncontrollably.
Lung cells do not normally respond to because they do not express the receptor; mutations resulting in a constitutively active receptor would not affect lung cells.
22. A normal tumor suppressor gene encodes a protein that restrains cell division. A mutant form of the protein fails to suppress cell division, but if either of the two alleles in an individual encodes a normal protein, normal function will continue. A normal oncogene encodes a regulator protein that triggers cell division, but only when an appropriate signal (growth factor) is present. The mutant version of the oncogene product constantly sends the signal to divide, whether or not growth factors are present.
23. In a child who develops multiple tumors in both eyes, every retinal cell had a defective copy of the Rb gene at birth. Early in the child’s life, several cells independently underwent a second mutation that damaged the one good Rb allele, producing a tumor. A child who develops a single tumor had, at birth, two good copies of the Rb gene in every cell; mutation in both Rb alleles in one cell (extremely rare) caused the single tumor.
24. Two cells expressing the same surface receptor may have access to different complements of target proteins for protein phosphorylation and therefore have different responses to the same signal.
25.
The data favor the cell-based model, which predicts different receptors present on different cells.
This experiment addresses the issue of the independence of different taste sensations. Even though the receptors for sweet and/or umami are missing, the animals’ other taste sensations are normal; thus, pleasant and unpleasant taste sensations are independent.
Yes. Loss of either T1R1 or T1R3 subunits abolishes umami taste sensation.
Both models. With either model, removing one receptor would abolish that taste sensation.
Yes. Loss of either the T1R2 or T1R3 subunits almost completely abolishes the sweet taste sensation; complete elimination of sweet taste requires deletion of both subunits.
At very high sucrose concentrations, T1R2 and, to a lesser extent, T1R3 receptors, as homodimers, can detect sweet taste.
The results are consistent with either model of taste encoding, but they do strengthen the researchers’ conclusions. Ligand binding can be completely separated from taste sensation. If the ligand for the receptor in “sweet-tasting cells” binds a molecule, mice prefer that molecule as a sweet compound.
Chapter 13
1. Consider the developing chick as the system; the nutrients, egg shell, and outside world are the surroundings. Transformation of the single cell into a chick drastically reduces the entropy of the system. Initially, the parts of the egg outside the embryo (the surroundings) contain complex fuel molecules (a low-entropy condition). During incubation, some of these complex molecules are converted to large numbers of and molecules (high entropy). This increase in the entropy of the surroundings is larger than the decrease in entropy of the chick (the system).
2.
7.56 kJ/mol
3.
262
608
0.30
4.
5.
6.
At a given temperature, the value of for any reaction is fixed and is defined for standard conditions (here, both fructose 6-phosphate and glucose 6-phosphate at 1 m). In contrast, ∆G is a variable that can be calculated for any set of reactant and product concentrations.
7.
8. Less. The overall equation for ATP hydrolysis can be approximated as
(This is only an approximation, because the ionized species shown here are the major, but not the only, forms present.) Under standard conditions , the concentration of water is 55 m and does not change during the reaction. Because ions are produced in the reaction, at a higher [] (pH 5.0) the equilibrium would be shifted to the left and less free energy would be released.
9. 10
10.
∆G for ATP hydrolysis is lower when [ATP]/[ADP] is low than when [ATP]/[ADP] is high. Less energy is available to the cell from a given [ATP] when the [ATP]/[ADP] ratio falls and more is available when it rises.
11.
; . No. The cellular [glucose 6-phosphate] is much greater than this, favoring the reverse reaction.
11 m. No. The maximum solubility of glucose is less than 1 m.
651 . Yes. This reaction path can occur with a concentration of glucose that is readily soluble and does not produce a large osmotic force.
No. This would require such high that the phosphate salts of divalent cations would precipitate.
By directly transferring the phosphoryl group from ATP to glucose, the phosphoryl group transfer potential (“tendency” or “pressure”) of ATP is utilized without generating high concentrations of intermediates. The essential part of this transfer is, of course, the enzymatic catalysis.
12.
13.
68.7
14.
15. 46.7 kJ/mol
16. Isomerization moves the carbonyl group from C-1 to C-2, setting up a carbon–carbon bond cleavage between C-3 and C-4. Without isomerization, bond cleavage would occur between C-2 and C-3, generating one two-carbon compound and one four-carbon compound.
17. The mechanism is the same as that of the alcohol dehydrogenase reaction (see Fig. 14-12).
18. The first step is the reverse of an aldol condensation (see the aldolase mechanism, Fig. 14-5); the second step is an aldol condensation (see Fig. 13-4).
19.
Oxidation-reduction, dehydrogenase with NAD cofactor; also produced
Isomerization, isomerase
Internal rearrangement, isomerase
Phosphoryl group transfer, kinase and ATP; ADP produced
Hydrolysis, protease or peptidase and
Oxidation-reduction, dehydrogenase with NAD cofactor; also produced
Oxidation-reduction, dehydrogenase with NAD cofactor and ; also produced
20. ATP; the products of phosphoarginine hydrolysis are stabilized by resonance forms not available in the intact molecule.
21. Yes. If [ADP] and [polyphosphate] are kept high, and [ATP] is kept low, the actual free-energy change would be negative.
22.
46 kJ/mol
46 kg; 68%
ATP is synthesized as it is needed, then broken down to ADP and ; its concentration is maintained in a steady state.
23. The ATP system is in a dynamic steady state; [ATP] remains constant because the rate of ATP consumption equals its rate of synthesis. ATP consumption involves release of the terminal (γ) phosphoryl group; synthesis of ATP from ADP involves replacement of this group. Hence the terminal phosphoryl undergoes rapid turnover. In contrast, the central (β) phosphoryl undergoes only relatively slow turnover.
24.
1.7 kJ/mol
Inorganic pyrophosphatase catalyzes the hydrolysis of pyrophosphate and drives the net reaction toward the synthesis of acetyl-CoA.
25. Although all the options are possible in principle, the production of AMP and in the reaction tells us that AMP is the activating group.
26. 36 kJ/mol
27. (d) (a) (c) (b)
28.
Pyruvate/lactate
Lactate formation
29.
Initially, electrons will be given up by lactate (converting it to pyruvate) and will flow to fumarate, converting it to succinate.
The four reactants have reached their equilibrium concentrations; .
30.
1.14 V
~4
31.
32. In order of increasing tendency: (a), (d), (b), (c)
33. (c) and (d)
34.
0.0293
308
Q is much lower than , indicating that the PFK-1 reaction is far from equilibrium in cells; this reaction is slower than the subsequent reactions in glycolysis. Flux through the glycolytic pathway is largely determined by the activity of PFK-1.
35.
The physiological concentration (0.023 mm) is 16,000 times the equilibrium concentration; this reaction does not reach equilibrium in the cell. Many reactions in the cell are not at equilibrium.
36. Malate synthase is saturated with the substrate acetyl-CoA; its concentration is almost greater than for acetyl-CoA. But we are not given the concentration or of its other substrate (glyoxylate). If [glyoxylate] is below the for glyoxylate, the reaction rate is limited by [glyoxylate], and malate synthase is not operating at .
37.
The lowest-energy, highest-entropy state occurs when the dye concentration is the same in both cells. If a “fish trap” gap junction allowed unidirectional transport, more of the dye would end up in the oligodendrocyte and less in the astrocyte. This would be a higher-energy, lower-entropy state than the starting state, violating the second law of thermodynamics. The model proposed by Robinson et al. requires an impossible spontaneous change from a lower-energy state to a higher-energy state without an energy input—again, thermodynamically impossible.
Molecules, unlike fish, do not exhibit directed behavior; they move randomly by Brownian motion. Diffusion results in net movement of molecules from a region of higher concentration to a region of lower concentration simply because it is more likely that a molecule on the high-concentration side will enter the connecting channel. The narrower end, like the rate-limiting step of a metabolic pathway, limits the rate at which molecules pass through; random motion of the molecules is less likely to move them through the smaller cross section. The wide end of the channel does not act like a funnel for molecules because the narrow end limits the rate of movement equally in both directions. When the concentrations on both sides are equal, the rates of movement in both directions are equal and there will be no change in concentration.
Fish exhibit nonrandom behavior. Fish behavior favors forward movement and avoids crowding and narrow places. Fish that enter the large opening of the channel tend to move forward but then are unlikely to enter the small opening because of their preferred behavior.
Here are two of many possible explanations: (1) The dye could bind to a molecule in the oligodendrocyte. Binding effectively removes the dye from the bulk solvent, yet it remains visible in the fluorescence microscope. (2) The dye could be sequestered in a subcellular organelle of the oligodendrocyte, either actively pumped in at the expense of ATP or drawn in by its attraction to other molecules in that organelle.
Chapter 14
1. At equilibrium, In living cells, . The reaction is therefore far from equilibrium: the cellular concentrations of the products (glucose 6-phosphate and ADP) are much lower than expected at equilibrium, and those of the reactants are much higher. The reaction therefore tends strongly to go to the right.
2.
3.
4.
5. C-1. This experiment demonstrates the reversibility of the aldolase reaction. The C-1 of glyceraldehyde 3-phosphate is equivalent to C-4 of fructose 1,6-bisphosphate (see Fig. 14-6). The starting glyceraldehyde 3-phosphate must have been labeled at C-1. The C-3 of dihydroxyacetone phosphate becomes labeled through the triose phosphate isomerase reaction, thus giving rise to fructose 1,6-bisphosphate labeled at C-3.
6. No. There would be no anaerobic production of ATP; aerobic ATP production would be diminished only slightly.
7. No. Lactate dehydrogenase is required to recycle from the NADH formed during the oxidation of glyceraldehyde 3-phosphate.
8. The transformation of glucose to lactate occurs when myocytes are low in oxygen, and it provides a means of generating ATP under -deficient conditions. Because lactate can be oxidized to pyruvate, glucose is not wasted; pyruvate is oxidized by aerobic reactions when becomes plentiful. This metabolic flexibility gives the organism a greater capacity to adapt to its environment.
9. The cell rapidly removes the 1,3-bisphosphoglycerate in a favorable subsequent step, catalyzed by phosphoglycerate kinase.
10.
3-Phosphoglycerate would be the product.
In the presence of arsenate, there is no net ATP synthesis under anaerobic conditions.
11.
Ethanol fermentation requires 2 mol of per mole of glucose.
Ethanol is the reduced product formed during reoxidation of NADH to , and is the byproduct of the conversion of pyruvate to ethanol. Yes. Pyruvate must be converted to ethanol to produce a continuous supply of for the oxidation of glyceraldehyde 3-phosphate. Fructose 1,6-bisphosphate accumulates; it is formed as an intermediate in glycolysis.
Arsenate replaces in the glyceraldehyde 3-phosphate dehydrogenase reaction to yield an acyl arsenate, which spontaneously hydrolyzes. This prevents formation of ATP, but 3-phosphoglycerate continues through the pathway.
12. Dietary niacin is used to synthesize . Oxidations carried out by are part of cyclic processes, with as electron carrier (reducing agent); one molecule of can oxidize many thousands of molecules of glucose, and thus the dietary requirement for the precursor vitamin (niacin) is relatively small.
13. Dihydroxyacetone (catalyzed by a dehydrogenase)
14. Galactokinase deficiency: galactose (less toxic); transferase deficiency: galactose 1-phosphate (more toxic)
15. Consumption of alcohol forces competition for between ethanol metabolism and gluconeogenesis. The problem is compounded by strenuous exercise and lack of food, because at these times the level of blood glucose is already low.
16.
The rapid increase in glycolysis; the rise in pyruvate and NADH results in a rise in lactate.
Lactate is transformed to glucose via pyruvate. This is a slower process because formation of pyruvate is limited by availability, the lactate dehydrogenase equilibrium is in favor of lactate, and conversion of pyruvate to glucose is energy-requiring.
The equilibrium for the lactate dehydrogenase reaction is in favor of lactate formation.
17. Lactate is transformed to glucose in the liver by gluconeogenesis (see Fig. 14-16). A defect in FBPase-1 would prevent entry of lactate into the gluconeogenic pathway in hepatocytes, causing lactate to accumulate in the blood.
18. In the absence of , the ATP needs are met by anaerobic glucose metabolism (fermentation to lactate). Because aerobic oxidation of glucose produces far more ATP than does fermentation, less glucose is needed to produce the same amount of ATP.
19.
There are two binding sites for ATP: a catalytic site and a regulatory site. Binding of ATP to a regulatory site inhibits PFK-1, by reducing or increasing for ATP at the catalytic site.
Glycolytic flux is reduced when ATP is plentiful.
The graph indicates that increased [ADP] suppresses the inhibition by ATP. Because the adenine nucleotide pool is fairly constant, consumption of ATP leads to an increase in [ADP]. The data show that the activity of PFK-1 may be regulated by the [ATP]/[ADP] ratio.
20. The phosphate group of glucose 6-phosphate is completely ionized at pH 7, giving the molecule an overall negative charge. Because membranes are generally impermeable to electrically charged molecules, glucose 6-phosphate cannot pass from the bloodstream into cells and hence cannot enter the glycolytic pathway and generate ATP. (This is why glucose, once phosphorylated, cannot escape from the cell.)
21.
22.
When aldolase splits glucose into two trioses phosphates, C-3 and C-4 of glucose become C-1 of the glyceraldehyde 3-phosphate that proceeds through glycolysis.
23. Fermentation releases energy, some conserved in the form of ATP but much of it dissipated as heat. Unless the fermenter contents are cooled, the temperature would become high enough to kill the microorganisms.
24. Soybeans and wheat contain starch, a polymer of glucose. The microorganisms break down starch to glucose, glucose to pyruvate via glycolysis, and — because the process is carried out in the absence of (i.e., it is a fermentation) — pyruvate to lactate and ethanol. If were present, pyruvate would be oxidized to acetyl-CoA, then to and . Some of the acetyl-CoA, however, would also be hydrolyzed to acetic acid (vinegar) in the presence of oxygen.
25. (a), (b), and (d) are glucogenic; (c) and (e) are not.
26.
In the pyruvate carboxylase reaction, is added to pyruvate, but PEP carboxykinase removes the same in the next step. Thus, is not (initially) incorporated into glucose.
27. 4 ATP equivalents per glucose molecule
28. Gluconeogenesis would be highly endergonic, and it would be impossible to separately regulate gluconeogenesis and glycolysis.
29. The cell “spends” 1 ATP and 1 GTP in converting pyruvate to PEP.
30. The proteins are degraded to amino acids and used for gluconeogenesis.
31. Succinate transforms to oxaloacetate, which passes into the cytosol and is converted to PEP by PEP carboxykinase. Two moles of PEP are then required to produce a mole of glucose by the route outlined in Fig. 14-16.
32. If the catabolic and anabolic pathways of glucose metabolism are operating simultaneously, unproductive cycling of ADP and ATP occurs, with extra consumption.
33. At the very least, accumulation of ribose 5-phosphate would tend to force this reaction in the reverse direction by mass action (see Eqn 13-4). It might also affect other metabolic reactions that involve ribose 5-phosphate as a substrate or product — such as the pathways of nucleotide synthesis.
34.
Ethanol tolerance is likely to involve many more genes, and thus the engineering would be a much more involved project.
l-Arabinose isomerase (the araA enzyme) converts an aldose to a ketose by moving the carbonyl of a nonphosphorylated sugar from C-1 to C-2. No analogous enzyme is discussed in this chapter; all the enzymes described here act on phosphorylated sugars. An enzyme that carries out a similar transformation with phosphorylated sugars is phosphohexose isomerase. l-Ribulokinase (araB) phosphorylates a sugar at C-5 by transferring the phosphate from ATP. Many such reactions are described in this chapter, including the hexokinase reaction. l-Ribulose 5-phosphate epimerase (araD) switches the and groups on a chiral carbon of a sugar. No analogous reaction is described in the chapter, but it is described in Chapter 20 (see Fig. 20-31).
The three ara enzymes would convert arabinose to xylulose 5-phosphate by the following pathway: Arabinose l-ribulose 5-phosphate xylulose 5-phosphate.
The arabinose is converted to xylulose 5-phosphate as in (c), which enters the pathway in Fig. 14-31a; the glucose 6-phosphate product is then fermented to ethanol and .
6 molecules of arabinose molecules of ATP are converted to 6 molecules of xylulose 5-phosphate, which feed into the pathway in Fig. 14-31a to yield 5 molecules of glucose 6-phosphate, each of which is fermented to yield 3 ATP (they enter as glucose 6-phosphate, not glucose) — 15 ATP in all. Overall, you would expect a yield of from the 6 arabinose molecules. The other products are 10 molecules of ethanol and 10 molecules of .
Given the lower ATP yield, for an amount of growth (i.e., of available ATP) equivalent to growth without the added genes, the engineered Z. mobilis must ferment more arabinose, and thus it produces more ethanol.
One way to allow the use of xylose would be to add the genes for two enzymes: an analog of the araD enzyme that converts xylose to ribose by switching the and on C-3, and an analog of the araB enzyme that phosphorylates ribose at C-5. The resulting ribose 5-phosphate would feed into the existing pathway.
Chapter 15
1. 11 s
2.
In muscle: Glycogen breakdown supplies energy (ATP) via glycolysis. Glycogen phosphorylase catalyzes the conversion of stored glycogen to glucose 1-phosphate, which is converted to glucose 6-phosphate, an intermediate in glycolysis. During strenuous activity, skeletal muscle requires large quantities of glucose 6-phosphate. In the liver: Glycogen breakdown maintains a steady level of blood glucose between meals (glucose 6-phosphate is converted to free glucose).
In actively working muscle, ATP flux requirements are very high and glucose 1-phosphate must be produced rapidly, requiring a high .
3.
b., c. The value of this ratio in the cell indicates that [glucose 1-phosphate] is far below the equilibrium value. The rate at which glucose 1-phosphate is removed (through entry into glycolysis) is greater than its rate of production (by the glycogen phosphorylase reaction), so metabolite flow is from glycogen to glucose 1-phosphate. The glycogen phosphorylase reaction is probably the regulatory step in glycogen breakdown.
4.
Increases
Decreases
Increases
5. Resting: [ATP] high; [AMP] low; [acetyl-CoA] and [citrate] intermediate. Running: [ATP] intermediate; [AMP] high; [acetyl-CoA] and [citrate] low. Glucose flux through glycolysis increases during the anaerobic sprint because (1) the ATP inhibition of glycogen phosphorylase and PFK-1 is partially relieved, (2) AMP stimulates both enzymes, and (3) lower citrate and acetyl-CoA levels relieve their inhibitory effects on PFK-1 and pyruvate kinase, respectively.
6. The migrating bird relies on the highly efficient aerobic oxidation of fats, rather than the anaerobic metabolism of glucose used by a sprinting rabbit. The bird reserves its muscle glycogen for short bursts of energy during emergencies.
7. Case A: (f), (3); Case B: (c), (3); Case C: (h), (4); Case D: (d), (6)
8.
(1) Adipose: fatty acid synthesis slower. (2) Muscle: glycolysis, fatty acid synthesis, and glycogen synthesis slower. (3) Liver: glycolysis faster; gluconeogenesis, glycogen synthesis, and fatty acid synthesis slower; pentose phosphate pathway unchanged.
(1) Adipose and (3) liver: fatty acid synthesis slower because lack of insulin results in inactive acetyl-CoA carboxylase, the first enzyme of fatty acid synthesis. Glycogen synthesis inhibited by cAMP-dependent phosphorylation (thus activation) of glycogen synthase. (2) Muscle: glycolysis slower because GLUT4 is inactive, so glucose uptake is inhibited. (3) Liver: glycolysis slower because the bifunctional PFK-2/FBPase-2 is converted to the form with active FBPase-2, decreasing [fructose 2,6-bisphosphate], which allosterically stimulates phosphofructokinase and inhibits FBPase-1; this also accounts for the stimulation of gluconeogenesis.
9.
Elevated
Elevated
Elevated
10.
PKA cannot be activated in response to glucagon or epinephrine, and glycogen phosphorylase is not activated.
PP1 remains active, allowing it to dephosphorylate glycogen synthase (activating it) and glycogen phosphorylase (inhibiting it).
Phosphorylase remains phosphorylated (active), increasing the breakdown of glycogen.
Gluconeogenesis cannot be stimulated when blood glucose is low, leading to dangerously low blood glucose during periods of fasting.
11. The drop in blood glucose triggers release of glucagon by the pancreas. In the liver, glucagon activates glycogen phosphorylase by stimulating its cAMP-dependent phosphorylation and stimulates gluconeogenesis by lowering [fructose 2,6-bisphosphate], thus stimulating FBPase-1.
12.
Reduced capacity to mobilize glycogen; lowered blood glucose between meals
Reduced capacity to lower blood glucose after a carbohydrate meal; elevated blood glucose
Reduced concentration of fructose 2,6-bisphosphate (F26BP) in liver, stimulating glycolysis and inhibiting gluconeogenesis
Reduced [F26BP], stimulating gluconeogenesis and inhibiting glycolysis
Increased uptake of fatty acids and glucose; increased oxidation of both
Increased conversion of pyruvate to acetyl-CoA; increased fatty acid synthesis
13.
Given that each particle contains about 55,000 glucose residues, the equivalent free glucose concentration would be . This would present a serious osmotic challenge for the cell! (Body fluids have a substantially lower osmolarity.)
The lower the number of branches, the lower the number of free ends available for glycogen phosphorylase activity, and the slower the rate of glucose release. With no branches, there would be just one site for phosphorylase to act.
The outer tier of the particle would be too crowded with glucose residues for the enzyme to gain access to cleave bonds and release glucose.
The number of chains doubles in each succeeding tier: tier 1 has one chain , tier 2 has two , tier 3 has four , and so on. Thus, for t tiers, the number of chains in the outermost tier, , is .
The total number of chains is . Each chain contains glucose molecules, so the total number of glucose molecules, , is .
Glycogen phosphorylase can release all but four of the glucose residues in a chain of length . Therefore, from each chain in the outer tier it can release glucose molecules. Given that there are chains in the outer tier, the number of glucose molecules the enzyme can release, , is .
The volume of a sphere is . In this case, r is the thickness of one tier times the number of tiers, or . Thus .
You can show algebraically that the value of that maximizes f is independent of t. Choosing :
5 64 635 64 1,232 2,111 6 64 762 128 1,760 3,547 7 64 889 192 2,421 4,512 8 64 1,016 256 3,230 5,154 9 64 1,143 320 4,201 5,572 10 64 1,270 384 5,350 5,834 11 64 1,397 448 6,692 5,986 12 64 1,524 512 8,240 6,060 13 64 1,651 576 10,011 6,079 14 64 1,778 640 12,019 6,059 15 64 1,905 704 14,279 6,011 16 64 2,032 768 16,806 5,943 Note: The optimum value of (i.e., at maximum f) is 13. In nature, varies from 12 to 14, which corresponds to f values very close to the optimum. If you choose another value for t, the numbers will differ but the optimal will still be 13.
Chapter 16
1.
Citrate synthase:
Aconitase:
Isocitrate dehydrogenase:
-Ketoglutarate dehydrogenase:
Succinyl-CoA synthetase:
Succinate dehydrogenase:
Fumarase:
Malate dehydrogenase:
b., c. CoA, condensation; none, isomerization; , oxidative decarboxylation; , CoA, and thiamine pyrophosphate, oxidative decarboxylation; CoA, substrate-level phosphorylation; FAD, oxidation; none, hydration; , oxidation
2.
3.
4. From the structural formulas, we see that the carbon-bound H/C ratio of hexanoate is higher than that of glucose . Hexanoate is more reduced and yields more energy on complete combustion to and .
5.
6. TPP: thiazolium ring adds to carbon of pyruvate, then stabilizes the resulting carbanion by acting as an electron sink. Lipoic acid: oxidizes pyruvate to level of acetate (acetyl-CoA) and activates acetate as a thioester. CoA-SH: activates acetate as thioester. FAD: oxidizes lipoic acid. .
7. Lack of TPP, caused by thiamine deficiency, inhibits pyruvate dehydrogenase; pyruvate accumulates.
8. Oxidative decarboxylation; or ; -ketoglutarate dehydrogenase reaction
9. Oxygen consumption is a measure of the activity of the first two stages of cellular respiration: glycolysis and the citric acid cycle. The addition of oxaloacetate or malate stimulates the citric acid cycle and thus stimulates respiration. The added oxaloacetate or malate serves a catalytic role: it is regenerated in the latter part of the citric acid cycle.
10.
28 molecules
11. ADP (or GDP), , CoA-SH, TPP, ; not lipoic acid, which is covalently attached to the isolated enzymes that use it
12. The flavin nucleotides, FMN and FAD, would not be synthesized. Because FAD is required in the citric acid cycle, flavin deficiency would strongly inhibit the cycle.
13. Oxaloacetate might be withdrawn for aspartate synthesis or for gluconeogenesis. Oxaloacetate is replenished by the anaplerotic reactions catalyzed by PEP carboxykinase, PEP carboxylase, malic enzyme, or pyruvate carboxylase (see Fig. 16-15).
14. The terminal phosphoryl group of GTP can be transferred to ADP in a reaction catalyzed by nucleoside diphosphate kinase, with an equilibrium constant of 1.0: .
15.
Malonate is a competitive inhibitor of succinate dehydrogenase.
A block in the citric acid cycle stops NADH formation, which stops electron transfer, which stops respiration.
A large excess of succinate (substrate) overcomes the competitive inhibition.
16.
Add uniformly labeled and check for the release of .
Equally distributed in C-2 and C-3 of oxaloacetate; an infinite number of turns
17.
C-1
C-3
C-3
C-2 (methyl group)
C-4
C-4
Equally distributed in C-2 and C-3
18. Thiamine is required for the synthesis of TPP, a prosthetic group in the pyruvate dehydrogenase and -ketoglutarate dehydrogenase complexes. A thiamine deficiency reduces the activity of these enzyme complexes and causes the observed accumulation of precursors.
19. No. For every two carbons that enter as acetate, two leave the cycle as ; thus there is no net synthesis of oxaloacetate. Net synthesis of oxaloacetate occurs by the carboxylation of pyruvate, an anaplerotic reaction.
20. Yes. The citric acid cycle would be inhibited. Oxaloacetate is present at relatively low concentrations in mitochondria, and removing it for gluconeogenesis would tend to shift the equilibrium for the citrate synthase reaction toward oxaloacetate.
21.
Inhibition of aconitase
Fluorocitrate; competes with citrate; by a large excess of citrate
Citrate and fluorocitrate are inhibitors of PFK-1.
All catabolic processes necessary for ATP production are shut down.
22. Glycolysis:
Pyruvate carboxylase reaction:
Malate dehydrogenase reaction:
This sequence recycles nicotinamide coenzymes under anaerobic conditions. The overall reaction is . Four are produced per glucose, increasing the acidity and thus the tartness of the wine.
23.
24. The cycle participates in catabolic and anabolic processes. For example, it generates ATP by substrate oxidation, but also provides precursors for amino acid synthesis (see Fig. 16-15).
25.
Decreases
Increases
Decreases
26.
Citrate is produced through the action of citrate synthase on oxaloacetate and acetyl-CoA. Citrate synthase can be used for net synthesis of citrate when (1) there is a continuous influx of new oxaloacetate and acetyl-CoA and (2) isocitrate synthesis is restricted, as in a culture medium low in . Aconitase requires , so an -restricted medium restricts the synthesis of aconitase.
The overall reaction is
The overall reaction consumes . Because the cellular pool of this oxidized coenzyme is limited, it must be regenerated from NADH by the electron-transfer chain, with consumption of . Consequently, the overall conversion of sucrose to citric acid is an aerobic process and requires molecular oxygen.
27. Succinyl-CoA is an intermediate of the citric acid cycle; its accumulation signals reduced flux through the cycle, calling for reduced entry of acetyl-CoA into the cycle. Citrate synthase, by regulating the primary oxidative pathway of the cell, regulates the supply of NADH and thus the flow of electrons from NADH to .
28. Fatty acid catabolism increases [acetyl-CoA], which stimulates pyruvate carboxylase. The resulting increase in [oxaloacetate] stimulates acetyl-CoA consumption by the citric acid cycle, and [citrate] rises, inhibiting glycolysis at the level of PFK-1. In addition, increased [acetyl-CoA] inhibits the pyruvate dehydrogenase complex, slowing the utilization of pyruvate from glycolysis.
29. Oxygen is needed to recycle from the NADH produced by the oxidative reactions of the citric acid cycle. Reoxidation of NADH occurs during mitochondrial oxidative phosphorylation.
30. Increased inhibits the citric acid cycle by mass action at the three -reducing steps; high [NADH] shifts the equilibrium toward .
31. Toward citrate; for the citrate synthase reaction under these conditions is about .
32. Steps and are essential in the reoxidation of the enzyme’s reduced lipoamide cofactor.
33. Many answers are possible. A genetic defect in MPC1 or MPC2 switches pyruvate catabolism from the oxidative path (through acetyl-CoA and the citric acid cycle) to the anaerobic reduction of pyruvate to lactate, with a much-increased use of glucose for the glycolytic production of ATP. The increase in cytosolic lactate concentration would acidify that part of the cell. Citric acid cycle activity would slow or would draw on substrates other than glycolytic pyruvate, such as fatty acids from adipose tissue. Blood levels of pyruvate and lactate would rise and blood pH would drop, producing acidosis. Muscle would fatigue easily.
34. The citric acid cycle is so central to metabolism that a serious defect in any cycle enzyme would probably be lethal to the embryo.
35.
The only reaction in muscle tissue that consumes significant amounts of oxygen is cellular respiration, so consumption is a good proxy for respiration.
Freshly prepared muscle tissue contains some residual glucose; consumption is due to oxidation of this glucose.
Yes. Because the amount of consumed increased when citrate or 1-phosphoglycerol was added, both can serve as substrate for cellular respiration in this system.
Experiment I: Citrate is causing much more consumption than would be expected from its complete oxidation. Each molecule of citrate seems to be acting as though it were more than one molecule. The only possible explanation is that each molecule of citrate functions more than once in the reaction — which is how a catalyst operates. Experiment II: The key is to calculate the excess consumed by each sample compared with the control (sample 1).
Sample Substrate(s) added absorbed Excess consumed 1 No extra 342 0 2 0.3 mL 0.2 m 1-phosphoglycerol 757 415 3 0.15 mL 0.02 m citrate 431 89 4 0.3 mL 0.2 m 1-phosphoglycerol 1,385 1,043 If both citrate and 1-phosphoglycerol were simply substrates for the reaction, you would expect the excess consumption by sample 4 to be the sum of the individual excess consumptions by samples 2 and 3 . However, the excess consumption when both substrates are present is roughly twice this amount . Thus citrate increases the ability of the tissue to metabolize 1-phosphoglycerol. This behavior is typical of a catalyst. Both experiments (I and II) are required to make this case convincing. Based on experiment I only, citrate is somehow accelerating the reaction, but it is not clear whether it acts by helping substrate metabolism or by some other mechanism. Based on experiment II only, it is not clear which molecule is the catalyst, citrate or 1-phosphoglycerol. Together, the experiments show that citrate is acting as a “catalyst” for the oxidation of 1-phosphoglycerol.
Given that the pathway can consume citrate (see sample 3), if citrate is to act as a catalyst it must be regenerated. If the set of reactions first consumes then regenerates citrate, it must be a circular rather than a linear pathway.
When the pathway is blocked at -ketoglutarate dehydrogenase, citrate is converted to -ketoglutarate but the pathway goes no further. Oxygen is consumed by reoxidation of the NADH produced by isocitrate dehydrogenase.
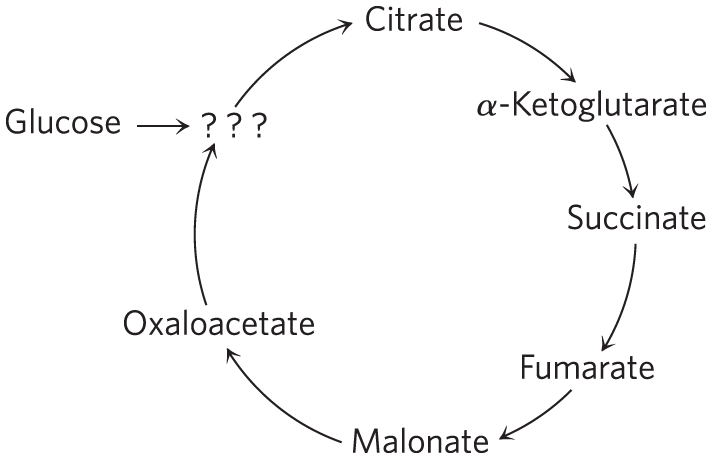
This differs from Fig. 16-7 in that it does not include cis-aconitate and isocitrate (between citrate and -ketoglutarate), or succinyl-CoA, or acetyl-CoA.
Establishing a quantitative conversion was essential to rule out a branched or other, more complex pathway.








Chapter 17
1. The fatty acid portion; the carbons in fatty acids are more reduced than those in glycerol.
2. Response to glucagon or epinephrine would be prolonged, mobilizing more fatty acids in adipocytes.
3. Fatty acyl groups condensed with CoA in the cytosol are first transferred to carnitine, releasing CoA, then transported into the mitochondrion, where they are again condensed with CoA. The cytosolic and mitochondrial pools of CoA are thus kept separate, and no radioactive CoA from the cytosolic pool enters the mitochondrion.
4. Malonyl-CoA would no longer inhibit entry of fatty acids into the mitochondrion for oxidation, so there might be a futile cycle of simultaneous fatty acid synthesis in the cytosol and fatty acid breakdown in mitochondria.
5.
The carnitine-mediated entry of fatty acids into mitochondria is the rate-limiting step in fatty acid oxidation. Carnitine deficiency slows fatty acid oxidation; added carnitine increases the rate.
All increase the metabolic need for fatty acid oxidation.
Carnitine deficiency might result from a deficiency of a carnitine precursor (such as lysine), or from a defect in one of the enzymes in the biosynthesis of carnitine.
6.
48 days
0.48 lb/day
7. The first step in fatty acid oxidation is analogous to the conversion of succinate to fumarate; the second step, to the conversion of fumarate to malate; the third step, to the conversion of malate to oxaloacetate.
8. 8 cycles; the last releases 2 acetyl-CoA.
9.
Irreversible hydrolysis of to by cellular inorganic pyrophosphatase
10. -Dodecanoyl-CoA; it is converted to -dodecanoyl-CoA, then -hydroxydodecanoyl-CoA.
11. 4 acetyl-CoA and 1 propionyl-CoA
12. Yes. Some of the tritium is removed from palmitate during the dehydrogenation reactions of oxidation. The removed tritium appears as tritiated water.
13.
In the pigeon, oxidation predominates; in the pheasant, anaerobic glycolysis of glycogen predominates.
Pigeon muscle would consume more .
Fat contains more energy per gram than glycogen does. In addition, the anaerobic breakdown of glycogen is limited by the tissue’s tolerance to lactate buildup. Thus the pigeon, using the oxidative catabolism of fats, is the long-distance flyer.
The enzymes listed in the table are the regulatory enzymes of their respective pathways and thus limit ATP production rate; however, triose phosphate isomerase and malate dehydrogenase are not regulatory enzymes of their respective pathways.
14. Oxidation of fats releases metabolic water; 1.5 L of water per kg of tripalmitoylglycerol (ignores the small contribution of glycerol to the mass).
15.
; phenylacetic acid
Even; removal of two carbons at a time from odd-number chains would leave phenylpropionate.
16. Because the mitochondrial pool of CoA is small, CoA must be recycled from acetyl-CoA via the formation of ketone bodies. This allows the operation of the -oxidation pathway, necessary for energy production.
17.
Glucose yields pyruvate via glycolysis, and pyruvate is the main source of oxaloacetate. Without glucose in the diet, [oxaloacetate] drops and the citric acid cycle slows.
Odd-number; propionate conversion to succinyl-CoA provides intermediates for the citric acid cycle and four-carbon precursors for gluconeogenesis.
18. For the odd-number heptanoic acid, oxidation produces propionyl-CoA, which can be converted in several steps to oxaloacetate, a starting material for gluconeogenesis. The even-number fatty acid cannot support gluconeogenesis, because it is entirely oxidized to acetyl-CoA.
19. Oxidation of -fluorooleate forms fluoroacetyl-CoA, which enters the citric acid cycle and produces fluorocitrate, a powerful inhibitor of aconitase. Inhibition of aconitase shuts down the citric acid cycle. Without reducing equivalents from the citric acid cycle, oxidative phosphorylation (ATP synthesis) is fatally slowed.
20. Ser to Ala: the Ala side chain in ACC cannot be phosphorylated (and thereby inactivated). Malonyl-CoA continues to be made, which inhibits carnitine acyltransferase 1. oxidation in mitochondria is blocked. Ser to Asp: ACC has a negative charge where Ser would normally be phosphorylated, so it remains inactive. Fatty acid synthesis is blocked and oxidation is stimulated (derepressed).
21. Enz-FAD, having a more positive standard reduction potential, is a better electron acceptor than , and the reaction is driven in the direction of fatty acyl–CoA oxidation. This more favorable equilibrium is obtained at the cost of 1 ATP; only 1.5 ATP are produced per oxidized in the respiratory chain (vs. 2.5 per NADH).
22. 9 turns; arachidic acid, a 20-carbon saturated fatty acid, yields 10 molecules of acetyl-CoA, the last two formed in the ninth turn.
23. See Fig. 17-12. is formed, which gives rise to oxaloacetate labeled at C-2 and C-3.
24. →→→ . All malate carbons would be labeled, but C-1 and C-4 would have only half as much label as C-2 and C-3.
25. ATP hydrolysis in the energy-requiring reactions of a cell takes up water in the reaction ; thus, in the steady state, there is no net production of .
26. Digestion of propionate requires methylmalonyl-CoA mutase, which, in turn, requires the cobalt-containing cofactor formed from vitamin .
27. Mass lost per day is about 0.66 kg, or about 140 kg in seven months. Ketosis could be avoided by degradation of nonessential body proteins to supply amino acid skeletons for gluconeogenesis.
28.
Fatty acids are converted to their CoA derivatives by enzymes in the cytoplasm; the acyl-CoAs are then imported into mitochondria for oxidation. Given that the researchers were using isolated mitochondria, they had to use CoA derivatives.
Stearoyl-CoA was rapidly converted to 9 acetyl-CoA by the -oxidation pathway. All intermediates reacted rapidly, and none were detectable at significant levels.
Two rounds. Each round removes two carbon atoms, thus two rounds convert an 18-carbon to a 14-carbon fatty acid and 2 acetyl-CoA.
The is higher for the trans isomer than for the cis, so a higher concentration of trans isomer is required for the same rate of breakdown. Roughly speaking, the trans isomer binds less well than the cis, probably because differences in shape, even though not at the target site for the enzyme, affect substrate binding to the enzyme.
The substrate for LCAD/VLCAD builds up differently, depending on the particular substrate; this is expected for the rate-limiting step in a pathway.
The kinetic parameters show that the trans isomer is a poorer substrate than the cis for LCAD, but there is little difference for VLCAD. Because it is a poorer substrate, the trans isomer accumulates to higher levels than the cis.
One possible pathway is shown below (indicating “inside” and “outside” mitochondria).
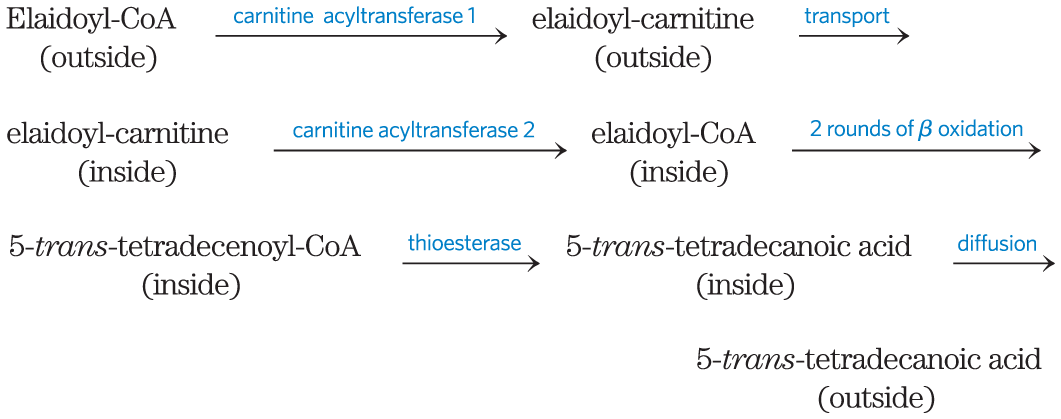
It is correct insofar as trans fats are broken down less efficiently than cis fats, and thus trans fats may “leak” out of mitochondria. It is incorrect to say that trans fats are not broken down by cells; they are broken down, but at a slower rate than cis fats.
Chapter 18
1.
2. This is a coupled-reaction assay. The product of the slow transamination (pyruvate) is rapidly consumed in the subsequent “indicator reaction” catalyzed by lactate dehydrogenase, which consumes NADH. Thus the rate of disappearance of NADH is a measure of the rate of the aminotransferase reaction. The indicator reaction is monitored by observing the decrease in absorption of NADH at 340 nm with a spectrophotometer.
3. Alanine and glutamine play special roles in the transport of amino groups from muscle and from other nonhepatic tissues, respectively, to the liver.
4. GTP is a product of the citric acid cycle, generated in the second step after the formation of -ketoglutarate. Elimination of the GTP inhibition of glutamate dehydrogenase leads to uncontrolled production of -ketoglutarate, which is oxidized to produce elevated levels of ATP. This in turn leads to insulin secretion.
5. No. The nitrogen in alanine can be transferred to oxaloacetate via transamination, to form aspartate.
6. 15 mol of ATP per mol of lactate; 13 mol of ATP per mol of alanine, when nitrogen removal is included
7.
Fasting resulted in low blood glucose; subsequent administration of the experimental diet led to rapid catabolism of glucogenic amino acids.
Oxidative deamination caused the rise in levels; the absence of arginine (an intermediate in the urea cycle) prevented conversion of to urea; arginine is not synthesized in sufficient quantities in the cat to meet the needs imposed by the stress of the experiment. This suggests that arginine is an essential amino acid in the cat’s diet.
Ornithine is converted to arginine by the urea cycle.
8.

9. The second amino group introduced into urea is transferred from aspartate, which is generated during the transamination of glutamate to oxaloacetate, a reaction catalyzed by aspartate aminotransferase. Approximately one-half of all the amino groups excreted as urea must pass through the aspartate aminotransferase reaction, making this the most highly active aminotransferase.
10.
A person on a diet consisting only of protein must use amino acids as the principal source of metabolic fuel. Because the catabolism of amino acids requires the removal of nitrogen as urea, the process consumes abnormally large quantities of water to dilute and excrete the urea in the urine. Furthermore, electrolytes in the “liquid protein” must be diluted with water and excreted. If the daily water loss through the kidney is not balanced by a sufficient water intake, a net loss of body water results.
When considering the nutritional benefits of protein, one must keep in mind the total amount of amino acids needed for protein synthesis and the distribution of amino acids in the dietary protein. Gelatin contains a nutritionally unbalanced distribution of amino acids. As large amounts of gelatin are ingested and the excess amino acids are catabolized, the capacity of the urea cycle may be exceeded, leading to ammonia toxicity. This is further complicated by the dehydration that may result from excretion of large quantities of urea. A combination of these two factors could produce coma and death.
11. Lysine and leucine
12.
Phenylalanine hydroxylase; a low-phenylalanine diet
The normal route of phenylalanine metabolism via hydroxylation to tyrosine is blocked, and phenylalanine accumulates.
Phenylalanine is transformed to phenylpyruvate by transamination, and then to phenyllactate by reduction. The transamination reaction has an equilibrium constant of 1.0, and phenylpyruvate is formed in significant amounts when phenylalanine accumulates.
Deficiency in production of tyrosine, which is a precursor of melanin, the pigment normally present in hair, leads to patches of unpigmented hair.
13. Not all amino acids are affected. Catabolism of the carbon skeletons of valine, methionine, and isoleucine is impaired because a functional methylmalonyl-CoA mutase (a coenzyme enzyme) is absent. The physiological effects of loss of this enzyme are described in Table 18-2 and Box 18-2.
14. The vegan diet lacks vitamin , leading to the increase in homocysteine and methylmalonate (reflecting the deficiencies in methionine synthase and methylmalonic acid mutase, respectively) in individuals on the diet for several years. Dairy products provide some vitamin in the lactovegetarian diet.
15. The genetic forms of pernicious anemia generally arise as a result of defects in the pathway that mediates absorption of dietary vitamin (see Box 17-2). Because dietary supplements are not absorbed in the intestine, these conditions are treated by injecting supplementary directly into the bloodstream.
16. The mechanism is identical to that for serine dehydratase (see Fig. 18-20a), except that the extra methyl group of threonine is retained, yielding -ketobutyrate instead of pyruvate.
17.
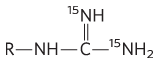
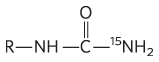
No label
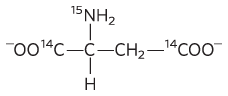
18.
Isoleucine
Step transamination, no analogous reaction, PLP; oxidative decarboxylation, analogous to the pyruvate dehydrogenase reaction, , TPP, lipoate, FAD; oxidation, analogous to the succinate dehydrogenase reaction, FAD; hydration, analogous to the fumarase reaction, no cofactor; oxidation, analogous to the malate dehydrogenase reaction, ; thiolysis (reverse aldol condensation), analogous to the thiolase reaction, CoA
19. A likely mechanism is
The formaldehyde (HCHO) produced in the second step reacts rapidly with tetrahydrofolate at the enzyme active site to produce -methylenetetrahydrofolate (see Fig. 18-17).
20.
Transamination; no analogies; PLP
Oxidative decarboxylation; analogous to oxidative decarboxylation of pyruvate to acetyl-CoA prior to entry into the citric acid cycle, and of -ketoglutarate to succinyl-CoA in the citric acid cycle; , FAD, lipoate, and TPP
Dehydrogenation (oxidation); analogous to dehydrogenation of succinate to fumarate in the citric acid cycle, and of fatty acyl–CoA to enoyl-CoA in oxidation; FAD
Carboxylation; no analogies in citric acid cycle or oxidation; ATP and biotin
Hydration; analogous to hydration of fumarate to malate in the citric acid cycle, and of enoyl-CoA to 3-hydroxyacyl-CoA in oxidation; no cofactors
Reverse aldol reaction; analogous to reverse of citrate synthase reaction in the citric acid cycle; no cofactors
21. Most amino acid catabolism occurs in the liver, including the key steps that are blocked in maple syrup urine disease. A liver transplant from a suitable donor with a normally functioning branched-chain -keto acid dehydrogenase complex could alleviate disease symptoms.
22.
Leucine; valine; isoleucine
Cysteine (derived from cystine). If cysteine were decarboxylated as shown in Fig. 18-6, it would yield , which could be oxidized to taurine.
The January 1957 blood shows significantly elevated levels of isoleucine, leucine, methionine, and valine; the January 1957 urine, significantly elevated isoleucine, leucine, taurine, and valine.
All patients had high levels of isoleucine, leucine, and valine in both blood and urine, suggesting a defect in the breakdown of these amino acids. Given that the urine also contained high levels of the keto forms of these three amino acids, the block in the pathway must occur after deamination but before dehydrogenation (as shown in Fig. 18-28).
The model does not explain the high levels of methionine in blood and taurine in urine. The high taurine levels may be due to the death of brain cells during the end stage of the disease. However, the reasons for high levels of methionine in blood are unclear; the pathway of methionine degradation is not linked with the degradation of branched-chain amino acids. Increased methionine could be a secondary effect of buildup of the other amino acids. It is important to keep in mind that the January 1957 samples were from an individual who was dying, so comparing blood and urine results with those of a healthy individual may not be appropriate.
The following information is needed (and was eventually obtained by other workers): (1) The dehydrogenase activity is significantly reduced or missing in individuals with maple syrup urine disease. (2) The disease is inherited as a single-gene defect. (3) The defect occurs in a gene encoding all or part of the dehydrogenase. (4) The genetic defect leads to production of inactive enzyme.
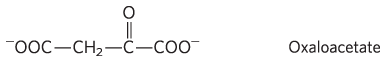
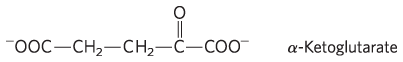

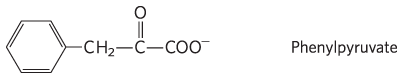
Chapter 19
1. Reaction 1: (a), (d) NADH; (b), (e) E-FMN; (c) and
Reaction 2: (a), (d) ; (b), (e) ; (c) and
Reaction 3: (a), (d) ; (b), (e) Q; (c) and
2. The side chain makes ubiquinone soluble in lipids and allows it to diffuse in the semifluid membrane.
3. From the difference in standard reduction potential for each pair of half-reactions, we can calculate . The oxidation of succinate by FAD is favored by the negative standard free-energy change . Oxidation by would require a large, positive, standard free-energy change .
4.
All carriers reduced; blocks the reduction of catalyzed by cytochrome oxidase.
All carriers reduced; in the absence of , the reduced carriers are not reoxidized.
All carriers oxidized
Early carriers more reduced; later carriers more oxidized
5.
Inhibition of NADH dehydrogenase by rotenone decreases the rate of electron flow through the respiratory chain, which in turn decreases the rate of ATP production. If this reduced rate is unable to meet the organism’s ATP requirements, the organism dies.
Antimycin A strongly inhibits the oxidation of Q in the respiratory chain, reducing the rate of electron transfer and leading to the consequences described in (a).
Because antimycin A blocks all electron flow to oxygen, it is a more potent poison than rotenone, which blocks electron flow from NADH but not from .
6.
The rate of electron transfer necessary to meet the ATP demand increases, and thus the P/O ratio decreases.
High concentrations of uncoupler produce P/O ratios near zero. The P/O ratio decreases and more fuel must be oxidized to generate the same amount of ATP. The extra heat released by this oxidation raises the body temperature.
Increased activity of the respiratory chain in the presence of an uncoupler requires the degradation of additional fuel. By oxidizing more fuel (including fat reserves) to produce the same amount of ATP, the body loses weight. When the P/O ratio approaches zero, the lack of ATP results in death.
7. Valinomycin acts as an uncoupler. It combines with to form a complex that passes through the inner mitochondrial membrane, dissipating the membrane potential. ATP synthesis decreases, which causes the rate of electron transfer to increase. This results in an increase in the gradient, consumption, and amount of heat released.
8. The steady-state concentration of in the cell is much higher than that of ADP. The released by ATP hydrolysis changes total very little.
9. Superoxide dismutase catalyzes the reduction of superoxide to hydrogen peroxide. The hydrogen peroxide can then be eliminated by glutathione peroxidase. This is a major pathway for eliminating the superoxide generated during respiration, helping to ameliorate the damaging effects of reactive oxygen species.
10.
External medium: ; matrix:
gradient contributes 1.7 kJ/mol toward ATP synthesis.
21
No
From the overall transmembrane potential
11.
5.5 s; to provide a constant level of ATP, regulation of ATP production must be tight and rapid.
12. . With a steady state [ATP] of , this is equivalent to 10 turnovers of the ATP pool per second; the reservoir would last about 0.13 s.
13. The citric acid cycle is stalled for lack of an acceptor of electrons from NADH. Pyruvate produced by glycolysis cannot enter the cycle as acetyl-CoA; accumulated pyruvate is transaminated to alanine and exported to the liver.
14. Cytosolic malate dehydrogenase plays a key role in the transport of reducing equivalents across the inner mitochondrial membrane via the malate-aspartate shuttle.
15. The inner mitochondrial membrane is impermeable to NADH, but the reducing equivalents of NADH are transferred (shuttled) through the membrane indirectly: they are transferred to oxaloacetate in the cytosol, the resulting malate is transported into the matrix, and mitochondrial is reduced to NADH.
16. Pyruvate dehydrogenase is located in mitochondria; glyceraldehyde 3-phosphate dehydrogenase, in the cytosol. The NAD pools are separated by the inner mitochondrial membrane.
17.
Glycolysis becomes anaerobic.
Oxygen consumption ceases.
Lactate formation increases.
ATP synthesis decreases to 2 ATP/glucose.
18. The response to (a), increased [ADP], is faster because the response to (b), reduced , requires protein synthesis.
19.
NADH is reoxidized via electron transfer instead of lactic acid fermentation.
Oxidative phosphorylation is more efficient.
The high mass-action ratio of the ATP system inhibits phosphofructokinase-1.
20. The Pasteur effect is not observed, because the citric acid cycle and respiratory chain are inactive. Fermentation to ethanol could be accomplished in the presence of , which is an advantage because strict anaerobic conditions are difficult to maintain.
21. Reactive oxygen species react with macromolecules, including DNA. If a mitochondrial defect leads to increased production of ROS, proto-oncogenes in the nuclear chromosomes can be damaged, producing oncogenes and leading to unregulated cell division and cancer (see Section 12.9).
22. Different extents of heteroplasmy for the defective gene produce different degrees of defective mitochondrial function.
23. Complete lack of glucokinase (two defective alleles) makes it impossible to carry out glycolysis at a sufficient rate to raise [ATP] to the threshold required for insulin secretion.
24. Defects in Complex II result in increased production of ROS, damage to DNA, and mutations that lead to unregulated cell division (cancer; see Section 12.9). It is not clear why the cancer tends to occur in the midgut.
25.
Unsaturated fatty acids increase membrane fluidity.
The cells cannot survive if their membranes contain less than 15% unsaturated fatty acids.
Oxygen is required for respiration. If cell growth is affected by membrane fluidity only in the presence of oxygen, then logically respiration rates may be affected by membrane fluidity.
The first observation indicates that something that migrates in the membrane is limiting the rate of respiration. The second observation suggests that respiration is inhibited when unsaturated fatty acid content is low. This might occur if membrane viscosity somehow affected the function of enzymes embedded in the membranes, the passive permeation of oxygen, or the rate of diffusion of a key factor in the membrane itself.
The overall conclusion is that ubiquinone diffusion through the membrane limits the rate of respiration.
Chapter 20
1. For the maximum photosynthetic rate, PSI (which absorbs light of 700 nm) and PSII (which absorbs light of 680 nm) must be operating simultaneously.
2. From water consumed in the overall reaction
3. is the hydrogen donor in photosynthesis. No is evolved, because is not split; the single photosystem lacks the water-splitting cofactor.
4.
Stops
Slows; some electron flow continues by the cyclic pathway.
5. During illumination, a proton gradient is established. When ADP and are added, ATP synthesis is driven by the gradient, which becomes exhausted in the absence of light.
6. DCMU blocks electron transfer between PSII and the first site of ATP production.
7. Venturicidin blocks proton movement through the complex; electron flow ( evolution) continues only until the free-energy cost of pumping protons against the rising proton gradient equals the free energy available in a photon. DNP, by dissipating the proton gradient, restores electron flow and evolution.
8. From the difference in reduction potentials, you can calculate for the redox reaction. Fig. 20-4 shows that the energy of photons in any region of the visible spectrum is more than sufficient to drive this endergonic reaction. Even photons in the infrared spectrum can provide sufficient energy.
9. ; the reaction is highly unfavorable! In chloroplasts, the input of light energy overcomes this barrier.
10.
11. No. The electrons from flow to the artificial electron acceptor , not to .
12. About once every 0.1 s; 1 in is excited
13. Light of 700 nm excites PSI, but not PSII; electrons flow from P700 to , but no electrons flow from P680 to replace them. When light of 680 nm excites PSII, electrons tend to flow to PSI, but the electron carriers between the two photosystems quickly become completely reduced.
14. No. The excited electron from P700 returns to refill the electron “hole” created by illumination. PSII is not needed to supply electrons, and no is evolved from . No NADPH is formed, because the excited electron returns to P700. Cyclic photophosphorylation produces ATP rather than NADPH.
15. ATP and NADPH are generated in the light and are essential for fixation; conversion stops as the supply of ATP and NADPH becomes exhausted. Some enzymes are switched off in the dark.
16. X is 3-phosphoglycerate; Y is ribulose 1,5-bisphosphate.
17. Ribulose 5-phosphate kinase, fructose 1,6-bisphosphatase, sedoheptulose 1,7-bisphosphatase, and glyceraldehyde 3-phosphate dehydrogenase; all are activated by reduction of a critical disulfide bond to a pair of sulfhydryls, which iodoacetate then blocks irreversibly.
18. The reductive pentose phosphate pathway regenerates ribulose 1,5-bisphosphate from triose phosphates produced during photosynthesis. The oxidative pentose phosphate pathway provides NADPH for reductive biosynthesis and ribose 5-phosphate for nucleotide synthesis.
19. Both types of “respiration” occur in plants, consume , and produce . (Mitochondrial respiration also occurs in animals.) Mitochondrial respiration takes place continuously, though primarily at night or on cloudy days; electrons derived from various fuels are passed through a chain of carriers in the inner mitochondrial membrane to . Photorespiration takes place in chloroplasts, peroxisomes, and mitochondria, during the daytime, when photosynthetic carbon fixation is occurring. Electron transfer in photorespiration is shown in Fig. 20-43; that for mitochondrial respiration, in Fig. 19-19.
20. In maize, the pathway fixes . Phosphoenolpyruvate (PEP) carboxylase carboxylates PEP to form oxaloacetate. Some of the oxaloacetate undergoes transamination to aspartate, but most undergoes reduction to malate in the mesophyll cells. Only after subsequent decarboxylation does the enter the Calvin cycle.
21. Measure the amount of fixed in leaves during an hour of darkness and an hour of bright illumination. The CAM plant will take up much more at night. Alternatively, measure the concentration of organic acids in the vacuoles by titrating an extract of leaves. In darkness, the plant will have a lower level of titratable acidity.
22. Isocitrate dehydrogenase reaction
23. Rates of photorespiration, which occurs when rubisco uses rather than as a substrate, are increased at higher light intensities and higher leaf temperatures. plants evolved mechanisms to minimize photorespiration, resulting in an increased ability to perform photosynthesis under these conditions. Because PEP carboxylase has a higher affinity for than rubisco, plants take up more under conditions of low . Thus, species 1 is a plant and species 2 is a plant.
24. is high in the cytosol because the cytosol lacks inorganic pyrophosphatase.
25.
Low in the cytosol and high [triose phosphate] in the chloroplast
High and [triose phosphate] in the cytosol
26. 3-Phosphoglycerate is the primary product of photosynthesis; rises when light-driven synthesis of ATP from ADP and slows.
27.
Fructose generated in the synthesis of dextran is readily imported and metabolized by the bacteria.
28. The first enzyme in each path is under reciprocal allosteric regulation. Inhibition of one path shunts isocitrate into the other path.
29.
(1) The presence of supports the hypothesis that chlorophyll is directly involved in catalysis of the phosphorylation reaction, . (2) Many enzymes (or other proteins) that contain are not phosphorylating enzymes, so the presence of in chlorophyll does not prove its role in phosphorylation reactions. (3) The presence of is essential to chlorophyll’s photochemical properties: light absorption and electron transfer.
(1) Enzymes catalyze reversible reactions, so an isolated enzyme that can, under certain lab conditions, catalyze removal of a phosphoryl group could probably, under different conditions (such as in cells), catalyze addition of a phosphoryl group. So, chlorophyll could be involved in the phosphorylation of ADP. (2) There are two possible explanations: the chlorophyll protein is a phosphatase only and does not catalyze ADP phosphorylation under cellular conditions, or the crude preparation contains a contaminating phosphatase activity that is unconnected to the photosynthetic reactions. (3) The preparation was probably contaminated with a nonphotosynthetic phosphatase activity.
(1) This light inhibition would be expected if the chlorophyll protein catalyzed the reaction . Without light, the reverse reaction, a dephosphorylation, would be favored. In the presence of light, energy is provided and the equilibrium would shift to the right, reducing the phosphatase activity. (2) This inhibition must be an artifact of the isolation or assay methods. (3) The crude preparation methods in use at the time were unlikely to preserve intact chloroplast membranes, so the inhibition must be an artifact.
In the presence of light, (1) ATP is synthesized and other phosphorylated intermediates are consumed; (2) glucose is produced and is metabolized by cellular respiration to produce ATP, with changes in the levels of phosphorylated intermediates; (3) ATP is produced and other phosphorylated intermediates are consumed.
Light energy is used to produce ATP (as in the Emerson model) and is used to produce reducing power (as in the Rabinowitch model).
The approximate stoichiometry for photophosphorylation is that 8 photons yield 2 NADPH and about 3 ATP. To reduce 1 requires 2 NADPH and 3 ATP. Thus, at a minimum, 8 photons are required per molecule reduced, in good agreement with Rabinowitch’s value.
Because the energy of light is used to produce both ATP and NADPH, each photon absorbed contributes more than just 1 ATP for photosynthesis. The process of energy extraction from light is more efficient than Rabinowitch supposed, and plenty of energy is available for this process — even with red light.
Chapter 21
1.
The 16 carbons of palmitate are derived from 8 acetyl groups of 8 acetyl-CoA molecules. The -labeled acetyl-CoA gives rise to malonyl-CoA labeled at C-1 and C-2.
The metabolic pool of malonyl-CoA, the source of all palmitate carbons except C-16 and C-15, does not become labeled with small amounts of -labeled acetyl-CoA. Hence, only [15,16-] palmitate is formed.
2. Both glucose and fructose are degraded to pyruvate in glycolysis. Pyruvate is converted to acetyl-CoA by the pyruvate dehydrogenase complex. Some of this acetyl-CoA enters the citric acid cycle, which produces reducing equivalents, NADH and NADPH. Mitochondrial electron transfer to yields ATP.
3.
4.
3 deuteriums per palmitate; all located on C-16; all other two-carbon units are derived from unlabeled malonyl-CoA
7 deuteriums per palmitate; located on all even-numbered carbons except C-16
5. By using the three-carbon unit malonyl-CoA, the activated form of acetyl-CoA (recall that malonyl-CoA synthesis requires ATP), metabolism is driven in the direction of fatty acid synthesis by the exergonic release of .
6.
The rate-limiting step in fatty acid synthesis is carboxylation of acetyl-CoA, catalyzed by acetyl-CoA carboxylase. High [citrate] and [isocitrate] indicate that conditions are favorable for fatty acid synthesis: an active citric acid cycle is providing a plentiful supply of ATP, reduced pyridine nucleotides, and acetyl-CoA. Citrate stimulates (increases the of) acetyl-CoA carboxylase
Because citrate binds more tightly to the filamentous (active) form of the enzyme, high [citrate] drives the protomer ⇌ filament equilibrium in the direction of the active form. In contrast, palmitoyl-CoA (the end product of fatty acid synthesis) drives the equilibrium in the direction of the inactive (protomer) form. Hence, when the end product of fatty acid synthesis accumulates, the biosynthetic path slows.
7.
1 ATP per acetyl group
Yes
8. No. The double bond in palmitoleate is introduced by an oxidation catalyzed by fatty acyl–CoA desaturase, a mixed-function oxidase that requires as a cosubstrate.
9.
10. In adult rats, stored triacylglycerols are maintained at a steady-state level through a balance of rates of degradation and biosynthesis. Hence, triacylglycerols of adipose (fat) tissue are constantly turned over, which explains the incorporation of label from dietary glucose.
11. Net reaction:
12. Methionine deficiency reduces the level of adoMet, which is required for de novo synthesis of phosphatidylcholine. The salvage pathway does not employ adoMet, but uses available choline. Thus phosphatidylcholine can be synthesized even when the diet is deficient in methionine, as long as choline is available.
13. During cholesterol biosynthesis, the two Claisen condensations involving acetyl-CoA and leading to HMG-CoA are both thermodynamically unfavorable. However, the product HMG-CoA is rapidly siphoned off by more thermodynamically favorable subsequent reactions. In fatty acid synthesis, all of the condensation reactions involving each new malonyl-CoA are identical. If acetyl-CoA were utilized instead of malonyl-CoA, all would be thermodynamically unfavorable. As subsequent reduction steps that expend NADPH in each four-step cycle are also thermodynamically unfavorable, there are no downstream processes capable of balancing the thermodynamics and pulling the sequence toward synthesis. The synthesis of long fatty acids would not be chemically feasible without the use of malonyl-CoA and the thermodynamic boost provided by decarboxylation.
14. label appears in three places in the activated isoprene:
15.
ATP
UDP-glucose
CDP-ethanolamine
UDP-galactose
Fatty acyl–CoA
S-Adenosylmethionine
Malonyl-CoA
-Isopentenyl pyrophosphate
16. Linoleate is required in the synthesis of prostaglandins. Animals cannot transform oleate to linoleate, so linoleate is an essential fatty acid. Plants can convert oleate to linoleate, and they provide animals with the required linoleate (see Fig. 21-12).
17. The rate-determining step in the biosynthesis of cholesterol is the synthesis of mevalonate, catalyzed by HMG-CoA reductase. This enzyme is allosterically regulated by mevalonate and cholesterol derivatives. High intracellular [cholesterol] also reduces transcription of the gene encoding HMG-CoA reductase.
18. When cholesterol levels decline because of treatment with a statin, cells attempt to compensate by increasing expression of the gene encoding HMG-CoA reductase; however, statins are good competitive inhibitors of HMG-CoA reductase activity and reduce overall production of cholesterol.
19. Note: In the absence of detailed knowledge of the literature on this enzyme, students might propose several plausible alternatives. Thiolase reaction: Begins with nucleophilic attack of an active-site Cys residue on the first acetyl-CoA substrate, displacing ―S-CoA and forming a covalent thioester link between Cys and the acetyl group. A base on the enzyme then extracts a proton from the methyl group of the second acetyl-CoA, leaving a carbanion that attacks the carbonyl carbon of the thioester formed in the first step. The sulfhydryl of the Cys residue is displaced, creating the product acetoacetyl-CoA. HMG-CoA synthase reaction: Begins in the same way, with a covalent thioester link formed between the enzyme’s Cys residue and the acetyl group of acetyl-CoA, with displacement of . The dissociates as CoA-SH, and acetoacetyl-CoA binds to the enzyme. A proton is abstracted from the methyl group of the enzyme-linked acetyl, forming a carbanion that attacks the ketone carbonyl of the acetoacetyl-CoA substrate. The carbonyl is converted to a hydroxyl ion in this reaction, which is protonated to create . The thioester link with the enzyme is then cleaved hydrolytically to generate the HMG-CoA product. HMG-CoA reductase reaction: Two successive hydride ions derived from NADPH first displace the , and then reduce the aldehyde to a hydroxyl group.
20. Statins inhibit HMG-CoA reductase, an enzyme in the pathway to the synthesis of activated isoprenes, which are precursors of cholesterol and a wide range of isoprenoids, including coenzyme Q (ubiquinone). Hence, statins might reduce the levels of coenzyme Q available for mitochondrial respiration. Ubiquinone is obtained in the diet as well as by direct biosynthesis, but it is not yet clear how much is required and how well dietary sources can substitute for reduced synthesis. Reductions in the levels of particular isoprenoids may account for some side effects of statins.
21.
Head-to-head. There are two ways to look at this. First, the “tail” of geranylgeranyl pyrophosphate has a branched dimethyl structure, as do both ends of phytoene. Second, no free is formed by the release of , indicating that the two “heads” are linked to form phytoene.
Four rounds of dehydrogenation convert four single bonds to double bonds.
No. A count of single and double bonds in the reaction below shows that one double bond is replaced by two single bonds—so, there is no net oxidation or reduction:
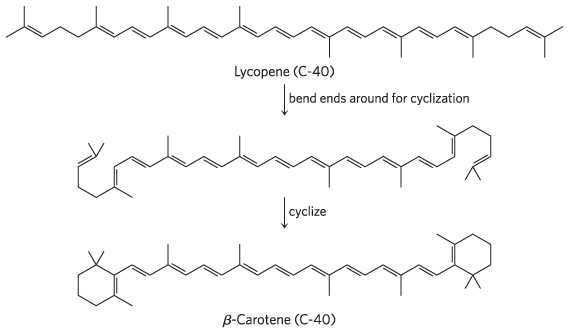
Steps through . The enzyme can convert IPP and DMAP to geranylgeranyl pyrophosphate, but it catalyzes no further reactions in the pathway, as confirmed by results with the other substrates.
Strains 1 through 4 lack crtE and have much lower astaxanthin production than strains 5 through 8, all of which overexpress crtE. Thus, overexpression of crtE leads to a substantial increase in astaxanthin production. Wild-type E. coli has some step activity, but this conversion of farnesyl pyrophosphate to geranylgeranyl pyrophosphate is strongly rate-limiting.
IPP isomerase. Comparing strains 5 and 6 shows that adding ispA, which catalyzes steps and , has little effect on astaxanthin production, so these steps are not rate-limiting. However, comparing strains 5 and 7 shows that adding idi substantially increases astaxanthin production, so IPP isomerase must be the rate-limiting step when crtE is overexpressed.
 “heads” are linked to form phytoene.
“heads” are linked to form phytoene.Chapter 22
1. In their symbiotic relationship with the plant, bacteria supply ammonium ion by reducing atmospheric nitrogen, a process that requires large quantities of ATP because of the very high activation energy of ammonia production from .
2. Fixed nitrogen is limiting in most environments, including marine ecosystems. Large increases in fixed nitrogen help to feed algae blooms, with an accompanying increase in aerobic respiration, which depletes oxygen in the affected waters.
3. A link is formed between enzyme-bound PLP and the phosphohomoserine substrate, with rearrangement to generate the ketimine at the carbon of the substrate. This activates the carbon for proton abstraction, leading to displacement of the phosphate and formation of a double bond between the and carbons. A rearrangement (beginning with proton abstraction at the pyridoxal carbon adjacent to the substrate amino nitrogen) moves the double bond and converts the ketimine to the aldimine form of PLP. Attack of water at the carbon is then facilitated by the linked pyridoxal, followed by hydrolysis of the imine link between PLP and the product, to generate threonine.
4. In the mammalian route, toxic ammonium ions are transformed to glutamine, reducing toxic effects on the brain.
5.
6. The amino-terminal glutaminase domain is similar in all glutamine amidotransferases. A drug that targeted this active site would probably inhibit many enzymes and produce many more side effects than a more specific inhibitor that targets the unique carboxyl-terminal synthetase active site.
7. If phenylalanine hydroxylase is defective, the biosynthetic route to tyrosine is blocked and tyrosine must be obtained from the diet.
8. The biosynthetic pathway requires reduction of the -carboxyl group of glutamate to a carbonyl. Prior acetylation of the -amino group prevents a spontaneous cyclization reaction that leads not to arginine, but to proline.
9. In adoMet synthesis, triphosphate is released from ATP. Hydrolysis of the triphosphate renders the reaction thermodynamically more favorable.
10. If the inhibition of glutamine synthase were not concerted, saturating concentrations of histidine would shut down the enzyme and cut off production of glutamine, which the bacterium needs to synthesize other products.
11. Folic acid is a precursor of tetrahydrofolate (see Fig. 18-16), required in the biosynthesis of glycine (see Fig. 22-14), a precursor of porphyrins. A folic acid deficiency therefore impairs hemoglobin synthesis.
12. This is a PLP-catalyzed decarboxylation.
13. Glycine auxotrophs: adenine and guanine; glutamine auxotrophs: adenine, guanine, and cytosine; aspartate auxotrophs: adenine, guanine, cytosine, and uridine
14. See Fig. 18-6, step , for the reaction mechanism of amino acid racemization. The F atom of fluoroalanine is an excellent leaving group. Fluoroalanine causes irreversible (covalent) inhibition of alanine racemase. One plausible mechanism (where Nuc denotes any nucleophilic amino acid side chain in the enzyme active site) is
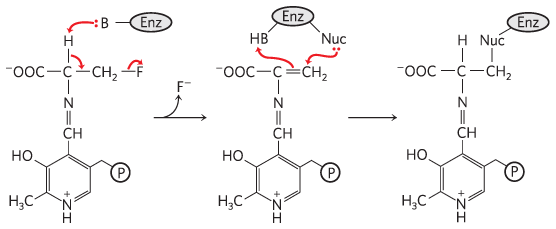
15.
As shown in Fig. 18-16, p-aminobenzoate is a component of tetrahydrofolate ( folate), the cofactor involved in the transfer of one-carbon units.
In the presence of sulfanilamide, a structural analog of p-aminobenzoate, bacteria are unable to synthesize tetrahydrofolate, a cofactor necessary for converting AICAR to FAICAR; thus, AICAR accumulates.
The competitive inhibition by sulfanilamide of the enzyme involved in tetrahydrofolate biosynthesis is overcome by the addition of excess substrate (p-aminobenzoate).
16. (b) and (d)
17. The -labeled orotate arises from the following pathway (the first three steps are part of the citric acid cycle):
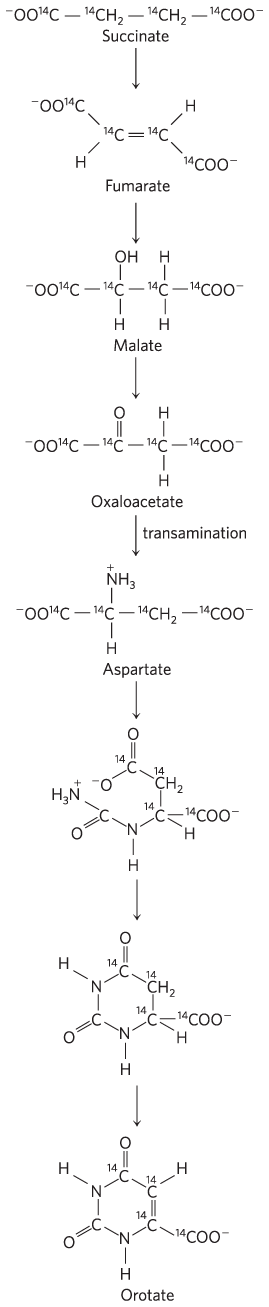
18. Organisms do not store nucleotides to be used as fuel, and they do not completely degrade them, but rather hydrolyze them to release the bases, which can be recovered in salvage pathways. The low C:N ratio of nucleotides makes them poor sources of energy.
19. Treatment with allopurinol has two consequences. (1) It inhibits conversion of hypoxanthine to uric acid, causing accumulation of hypoxanthine, which is more soluble and more readily excreted; this alleviates the clinical problems associated with AMP degradation. (2) It inhibits conversion of guanine to uric acid, causing accumulation of xanthine, which is less soluble than uric acid; this is the source of xanthine stones. Because the amount of GMP degradation is low relative to AMP degradation, the kidney damage caused by xanthine stones is less than the damage caused by untreated gout.
20. Tetrahydrofolate is utilized in thymidylate synthesis, and also in the synthesis of glycine from serine.
21.
The -carboxyl group is removed and an is added to the carbon.
BtrI has sequence homology with acyl carrier proteins. The molecular weight of BtrI increases when incubated under conditions in which CoA could be added to the protein. Adding CoA to a Ser residue would replace an , formula weight (FW) 17, with a -phosphopantetheine group (see Fig. 21-5; formula ), FW 356. Thus, , which is very close to the observed of 12,153.
The thioester could form with the -carboxyl group.
In the most common reaction for removing the -carboxyl group of an amino acid (see Fig. 18-6, reaction ), the carboxyl group must be free. Furthermore, it is difficult to imagine a decarboxylation reaction starting with a carboxyl group in its thioester form.
, close to the of (44). Given that BtrK is probably a decarboxylase, the most likely structure is the decarboxylated form:
. Glutamic acid , minus the (FW 17) removed in the glutamylation reaction, leaves a glutamyl group of FW 130; thus, -glutamylating the molecule shown above would add 130 to its . BtrJ is capable of -glutamylating other substrates, so it may -glutamylate this molecule. The most likely site for this is the free amino group, giving the following structure:
 ), the carboxyl group must be free. Furthermore, it is difficult to imagine a decarboxylation reaction starting with a carboxyl group in its thioester form.
), the carboxyl group must be free. Furthermore, it is difficult to imagine a decarboxylation reaction starting with a carboxyl group in its thioester form.Chapter 23
1. They are recognized by two different receptors, typically found in different cell types, and are coupled to different downstream effectors.
2. Ammonia is highly toxic to nervous tissue, especially the brain. In healthy individuals, excess is removed by transformation of glutamate to glutamine, which travels to the liver and is subsequently transformed to urea. The additional glutamine arises from conversion of glucose to -ketoglutarate, transamination of -ketoglutarate to glutamate, and conversion of glutamate to glutamine.
3. Glucogenic amino acids are used to make glucose for the brain; others are deaminated, then oxidized in mitochondria via the citric acid cycle.
4. From glucose, by the following route: Glucose → dihydroxyacetone phosphate (in glycolysis); dihydroxyacetone phosphate 3-phosphate (glycerol 3-phosphate dehydrogenase reaction)
5.
Increased muscular activity increases the demand for ATP, which is met by increased consumption.
After the sprint, lactate produced by anaerobic glycolysis is converted to glucose and glycogen, which requires ATP and therefore .
6. Glucose is the primary fuel for the brain. TPP-dependent oxidative decarboxylation of pyruvate to acetyl-CoA is essential to complete glucose metabolism. TPP synthesis requires the vitamin thiamine.
7. . For comparison, an Olympic pool, 50 m long, 25 m wide, and 2 m deep, holds .
8.
Inactivation provides a rapid means to change the concentration of active hormone and thus end its effects.
Changes in the rate of release from storage, the rate of conversion from prohormone to active hormone, and the rate of inactivation can rapidly change the level of a circulating peptide hormone.
9. Water-soluble hormones bind to receptors on the outer surface of the cell, triggering formation of a second messenger (e.g., cAMP) inside the cell. Lipid-soluble hormones can pass through the plasma membrane to act on target molecules or receptors directly.
10.
Skeletal muscle does not express glucose 6-phosphatase. Any glucose 6-phosphate produced enters the glycolytic pathway and, under -deficient conditions, is converted to lactate via pyruvate.
In a “fight-or-flight” situation, the concentration of glycolytic precursors must be high in preparation for muscular activity. Phosphorylated intermediates cannot escape from myocytes because the membrane is not permeable to charged species, and glucose 6-phosphate is not exported by the glucose transporter. In the liver, glucose is formed from glucose 6-phosphate and enters the bloodstream to maintain the blood glucose level in the homeostatic range.
11.
Excessive uptake and use of blood glucose by the liver, leading to hypoglycemia; shutdown of amino acid and fatty acid catabolism
Little circulating fuel is available for ATP requirements. Brain damage results because glucose is the main source of fuel for the brain.
12. As an uncoupler of oxidative phosphorylation, thyroxine would decrease the efficiency of the process (lower the P/O ratio), forcing the tissue to increase respiration to meet the normal demand for ATP. This less-efficient respiration would dissipate as heat a greater proportion of the energy potentially available for making ATP. Thermogenesis could also be due to the increased rate of ATP use by the thyroid-stimulated tissue. In this case, the efficiency of oxidative phosphorylation (the P/O ratio) would be unchanged. But because some energy is always dissipated as heat in the process, the increased production of ATP demanded by the stimulated tissue would produce more heat overall.
13. Because prohormones are inactive, they can be stored in quantity in secretory granules. Rapid activation is achieved by enzymatic cleavage in response to an appropriate signal.
14. In animals, glucose can be synthesized from many precursors (see Fig. 14-15). In humans, the principal precursors are glycerol from TAGs, glucogenic amino acids from protein degradation, and oxaloacetate formed by pyruvate carboxylase.
15. The ob/ob mouse, which is initially obese, will lose weight. The OB/OB mouse will retain its normal body weight.
16. . For a BMI of 25, weight must be 75 kg; he must lose 43 kg, or 95 lb.
17. Reduced insulin secretion. Valinomycin has the same effect as opening the channel, allowing exit and consequent hyperpolarization.
18. The liver does not receive the insulin message and therefore continues to have high levels of glucose 6-phosphatase and gluconeogenesis, increasing blood glucose both during a fast and after a glucose-containing meal. The elevated blood glucose triggers insulin release from pancreatic cells, hence the high level of insulin in the blood.
19. Some things to consider: What do the data show about the frequency of heart attack attributable to the drug among people taking the drug? How does this frequency compare with the data on individuals spared the long-term consequences of type 2 diabetes? Are other, equally effective treatment options with fewer adverse effects available?
20. Without intestinal glucosidase activity, absorption of glucose from dietary glycogen and starch is reduced, blunting the usual rise in blood glucose after a meal. The undigested oligosaccharides are fermented by intestinal bacteria, and the gases released cause intestinal discomfort.
21.
Increased; closing the ATP-gated channel would depolarize the membrane, increasing insulin release.
Type 2 diabetes results from decreased sensitivity to insulin, not a deficit of insulin production; increasing circulating insulin levels will reduce the symptoms associated with this disease.
Individuals with type 1 diabetes have deficient pancreatic cells, so glyburide will have no beneficial effect.
Iodine, like chlorine (the atom it replaces in the labeled glyburide), is a halogen, but it is a larger atom and has slightly different chemical properties. The iodinated glyburide might not bind to SUR. If it bound to another molecule instead, the experiment would result in cloning of the gene for this other, incorrect protein.
Although a protein has been “purified,” the “purified” preparation might be a mixture of several proteins that co-purify under those experimental conditions. In this case, the amino acid sequence could be that of a protein that co-purifies with SUR. Using antibody binding to show that the peptide sequences are present in SUR excludes this possibility.
Although the cloned gene does encode the 25 amino acid sequence found in SUR, it could be a gene that, coincidentally, encodes the same sequence in another protein. In this case, this other gene would most likely be expressed in different cells than the SUR gene. The mRNA hybridization results are consistent with the putative SUR cDNA actually encoding SUR.
The excess unlabeled glyburide competes with labeled glyburide for the binding site on SUR. As a result, there is significantly less binding of labeled glyburide, so little or no radioactivity is detected in the 140 kDa protein.
In the absence of excess unlabeled glyburide, labeled 140 kDa protein is found only in the presence of the putative SUR cDNA. Excess unlabeled glyburide competes with the labeled glyburide, and no -labeled 140 kDa protein is detected. This shows that the cDNA produces a glyburide-binding protein of the same molecular weight as SUR — strong evidence that the cloned gene encodes the SUR protein.
Several additional steps are possible, such as the following: (1) Express the putative SUR cDNA in CHO (Chinese hamster ovary) cells and show that the transformed cells have ATP-gated channel activity. (2) Show that HIT cells with mutations in the putative SUR gene lack ATP-gated channel activity. (3) Show that humans or experimental animals with mutations in the putative SUR gene are unable to secrete insulin.
Chapter 24
1. ; 290 times longer than the T2 phage head
2. The number of A residues does not equal the number of T residues, nor does the number of G equal the number of C, so the DNA is not a base-paired double helix; the M13 DNA is single-stranded.
3. ; ; ;
4. The exons contain 3 bp/amino amino . The remaining 864 bp are in introns, possibly in a leader or signal sequence, and/or in other noncoding DNA.
5. 5,000 bp.
Doesn’t change; Lk cannot change without breaking and re-forming the covalent backbone of the DNA.
Becomes undefined; a circular DNA with a break in one strand has, by definition, no Lk.
Decreases; in the presence of ATP, gyrase underwinds DNA.
Doesn’t change; this assumes that neither of the DNA strands is broken in the heating process.
6. For Lk to remain unchanged, the topoisomerase must introduce the same number of positive and negative supercoils.
7. ; probability
8.
Undefined; the strands of a nicked DNA could be separated and thus have no Lk.
476
The DNA is already relaxed, so the topoisomerase does not cause a net change; .
460; gyrase plus ATP reduces the Lk in increments of 2.
464; eukaryotic type I topoisomerases increase the Lk of underwound or negatively supercoiled DNA in increments of 1.
460; nucleosome binding does not break any DNA strands and thus cannot change Lk.
9. A fundamental structural unit in chromatin repeats about every 200 bp; the DNA is accessible to the nuclease only at 200 bp intervals. The brief treatment was insufficient to cleave the DNA at every accessible point, so a ladder of DNA bands is created in which the DNA fragments are multiples of 200 bp. The thickness of the DNA bands suggests that the distance between cleavage sites varies somewhat. For instance, not all the fragments in the lowest band are exactly 200 bp long.
10. A right-handed helix has a positive Lk; a left-handed helix (such as Z-DNA) has a negative Lk. Decreasing the Lk of a closed circular B-DNA by underwinding facilitates formation of regions of Z-DNA within certain sequences. (See Chapter 8, p. 273, for a description of sequences that permit the formation of Z-DNA.)
11.
Both strands must be covalently closed, and the molecule must be either circular or constrained at both ends.
Formation of cruciforms, left-handed Z-DNA, plectonemic or solenoidal supercoils, and unwinding of the DNA are favored.
E. coli DNA topoisomerase II or DNA gyrase
It binds the DNA at a point where it crosses on itself, cleaves both strands of one of the crossing segments, passes the other segment through the break, then reseals the break. The result is a change in Lk of .
12. Centromere, telomeres, and an autonomous replicating sequence or replication origin
13. The bacterial nucleoid is organized into domains approximately 10,000 bp long. Cleavage by a restriction enzyme relaxes the DNA within a domain, but not outside the domain. Any gene in the cleaved domain for which expression is affected by DNA topology will be affected by the cleavage; genes outside the domain will not.
14.
The lower, faster-migrating band is negatively supercoiled plasmid DNA. The upper band is nicked, relaxed DNA.
DNA topoisomerase I would relax the supercoiled DNA. The lower band would disappear, and all of the DNA would converge on the upper band.
DNA ligase would produce little change in the pattern. Some minor additional bands might appear near the upper band, due to the trapping of topoisomers not quite perfectly relaxed by the ligation reaction.
The upper band would disappear, and all of the DNA would be in the lower band. The supercoiled DNA in the lower band might become even more supercoiled and migrate somewhat faster.
15.
When DNA ends are sealed to create a relaxed, closed circle, some DNA species are completely relaxed but others are trapped in slightly underwound or overwound states. This gives rise to a distribution of topoisomers centered on the most relaxed species.
Positively supercoiled
The DNA that is relaxed despite the addition of dye is DNA with one or both strands broken. DNA isolation procedures inevitably introduce small numbers of strand breaks in some of the closed-circular molecules.
. This is determined by simply comparing native DNA with samples of known . In both gels, the native DNA migrates most closely with the sample of .
16. 62 million (the genome refers to the haploid genetic content of the cell; the cell is actually diploid, so the number of nucleosomes is doubled). The number is obtained by dividing 3.1 billion bp by 200 bp/nucleosome (giving 15.5 million nucleosomes), multiplying by 2 copies of H2A per nucleosome, and again multiplying by 2 to account for the diploid state of the cell. The 62 million would double upon replication.
17. DNA topoisomerase IV is needed to decatenate the two circular chromosome products of DNA replication prior to cell division.
18. A TAD, or topologically associating domain, is a DNA loop that is bound and constrained at its base. Supercoiling within the TAD is maintained in part by the restriction to free DNA rotation imposed by the protein binding at the base of the loop.
19.
In nondisjunction, one daughter cell and all of its descendants get two copies of the synthetic chromosome and are white; the other daughter cell and all of its descendants get no copies of the synthetic chromosome and are red. This gives rise to a half-white, half-red colony.
In chromosome loss, one daughter cell and all of its descendants get one copy of the synthetic chromosome and are pink; the other daughter cell and all of its descendants get no copies of the synthetic chromosome and are red. This gives rise to a half-pink, half-red colony.
The minimum functional centromere must be smaller than 0.63 kbp, because all fragments of this size or larger confer relative mitotic stability.
Telomeres are required to fully replicate only linear DNA; a circular molecule can replicate without them.
The larger the chromosome, the more faithfully it is segregated. The data show neither a minimum size below which the synthetic chromosome is completely unstable nor a maximum size above which stability no longer changes.
![A figure plots log [error rate (percent)] against length (k b p).](../images/8e_24ans_un05_329283.png)
As shown in the graph, even if the synthetic chromosomes were as long as the normal yeast chromosomes, they would not be as stable. This suggests that other, as yet undiscovered, elements are required for stability.
Chapter 25
1.
Structure 1



2. This is an extension of the classic Meselson-Stahl experiment. After three generations the molar ratio of DNA to DNA is .
3.
turns
40 min. In cells dividing every 20 min, a replicative cycle is initiated every 20 min, each cycle beginning before the prior one is complete.
2,000 to 5,000 Okazaki fragments. The fragments are 1,000 to 2,000 nucleotides long. The ligation of Okazaki fragments does not occur randomly. Each fragment is stably base-paired with the lagging strand template prior to ligation with its neighbor, ensuring proper ordering.
4. A, 28.7%; G, 21.3%; C, 21.3%; T, 28.7%. The DNA strand made from the template strand: A, 32.7%; G, 18.5%; C, 24.1%; T, 24.7%; the DNA strand made from the complementary template strand: A, 24.7%; G, 24.1%; C, 18.5%; T, 32.7%. This assumes that the two template strands are replicated completely.
5.
No. Incorporation of into DNA results from the synthesis of new DNA, which requires the presence of all four nucleotide precursors.
Yes. Although all four nucleotide precursors must be present for DNA synthesis, only one of them has to be radioactive for radioactivity to appear in the new DNA.
No. Radioactivity is incorporated only if the label is in the phosphate; DNA polymerase cleaves off pyrophosphate—that is, the - and -phosphate groups.
6. Mechanism 1: -OH group of an incoming dNTP attacks the phosphate of the triphosphate at the end of the growing DNA strand, displacing pyrophosphate. This mechanism uses normal dNTPs, and the growing end of the DNA always has a triphosphate on the end.
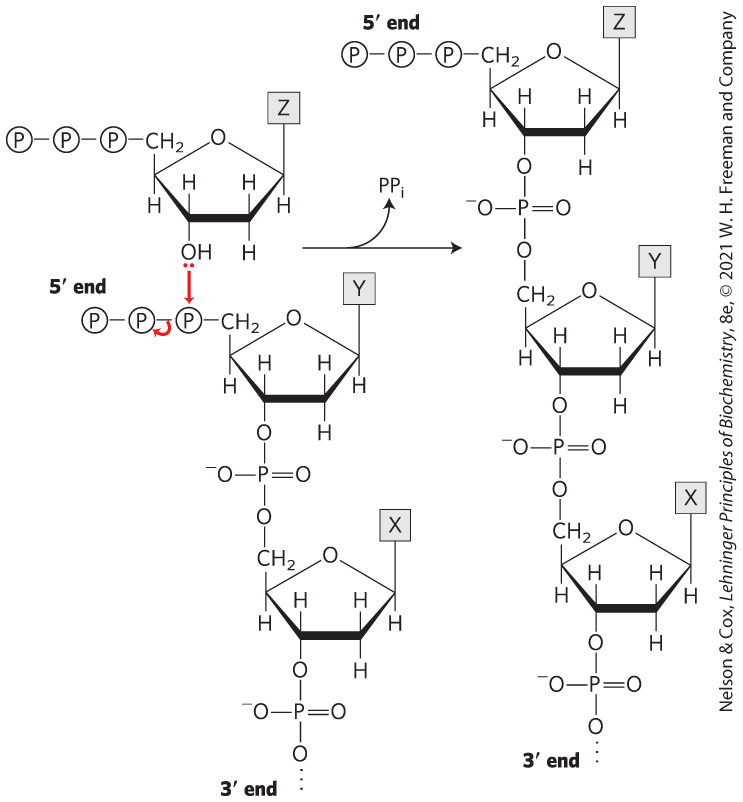
Mechanism 2: This uses a new type of precursor, nucleotide -triphosphates. The growing end of the DNA strand has a -OH group, which attacks the phosphate of an incoming deoxynucleoside -triphosphate, displacing pyrophosphate. Note that this mechanism would require the evolution of new metabolic pathways to supply the needed deoxynucleoside -triphosphates.
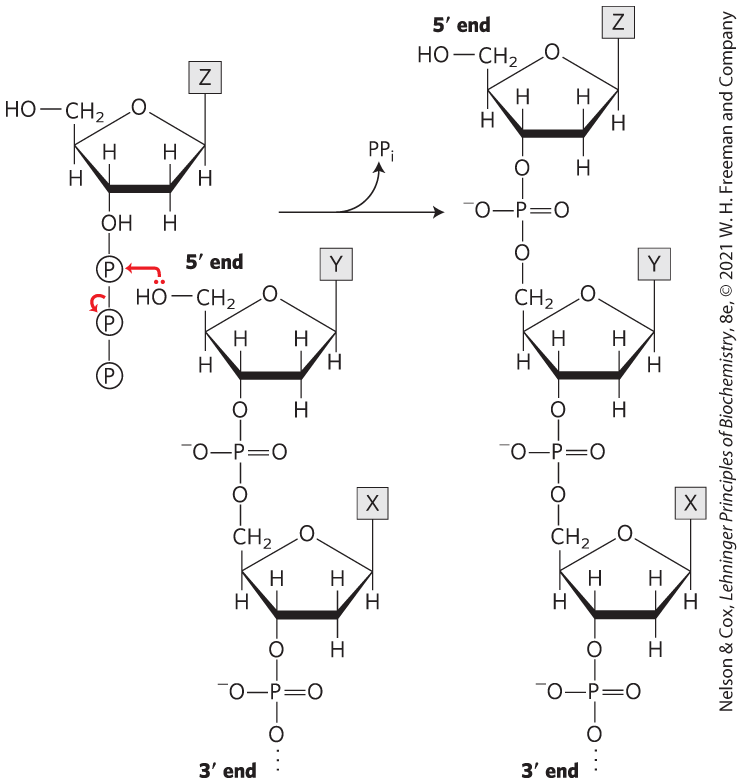
7. The DNA polymerase contains a exonuclease activity that degrades DNA to produce . The activity is not a exonuclease, because the addition of unlabeled dNTPs inhibits the production of (polymerization activity would suppress a proofreading exonuclease but not an exonuclease operating downstream of the polymerase). Addition of pyrophosphate would generate through reversal of the polymerase reaction.
8. Leading strand: Precursors: dATP, dGTP, dCTP, dTTP (also needs a template DNA strand and DNA primer); enzymes and other proteins: DNA gyrase, helicase, single-stranded DNA–binding protein, DNA polymerase III, topoisomerases, and pyrophosphatase. Lagging strand: Precursors: ATP, GTP, CTP, UTP, dATP, dGTP, dCTP, dTTP (also needs an RNA primer); enzymes and other proteins: DNA gyrase, helicase, single-stranded DNA–binding protein, primase, DNA polymerase III, DNA polymerase I, DNA ligase, topoisomerases, and pyrophosphatase. is also required as a cofactor for DNA ligase.
9. Mutants with defective DNA ligase produce a DNA duplex in which one of the strands remains in pieces (as Okazaki fragments). When this duplex is denatured, sedimentation results in one fraction containing the intact single strand (the high molecular weight band) and one fraction containing the unspliced fragments (the low molecular weight band).
10. Watson-Crick base pairing between template and leading strand; proofreading and removal of wrongly inserted nucleotides by the -exonuclease activity of DNA polymerase III. Yes — perhaps. Because the factors ensuring fidelity of replication are operative in both the leading and the lagging strands, the lagging strand would probably be made with the same fidelity. However, the greater number of distinct chemical operations involved in making the lagging strand might provide a greater opportunity for errors to arise.
11. bp (600 in each direction)
12. A small fraction (13 of cells) of the histidine-requiring mutants spontaneously undergo back-mutation and regain their capacity to synthesize histidine. 2-Aminoanthracene increases the rate of back-mutations about 1,800-fold and is therefore mutagenic. Since most carcinogens are mutagenic, 2-aminoanthracene is probably carcinogenic.
13. Spontaneous deamination of 5-methylcytosine (see Fig. 8-29a) produces thymine, and thus a G–T mismatched pair. These are among the most common mismatches in the DNA of eukaryotes. The specialized repair system restores the pair.
14. ∼1,950 (650 in the DNA degraded between the mismatch and GATC, plus 650 in DNA synthesis to fill the resulting gap, plus 650 in degradation of the pyrophosphate products to inorganic phosphate). ATP is hydrolyzed by the MutSL complex and by the UvrD helicase.
15.
UV irradiation produces pyrimidine dimers; in normal fibroblasts, these are excised by cleavage of the damaged strand by a special excinuclease. Thus the denatured single-stranded DNA contains the many fragments created by the cleavage, and the average molecular weight is lowered. These fragments of single-stranded DNA are absent from the XPG samples, as indicated by the unchanged average molecular weight.
The absence of fragments in the single-stranded DNA from the XPG cells after irradiation suggests the special excinuclease is defective or missing.
16. Most cancerous tumors consist of cells that are deficient in some aspect of DNA repair, relative to the normal surrounding tissue. They thus can be more sensitive to the DNA damaging agent. Tumor cells also tend to be actively dividing, a state in which cells are more sensitive to DNA damage that might be encountered by replication forks.
17. Using G* to represent :
TTG T ACGTG
TTGCACGTG
TTGCACGTG
TTGCACGTG
TTG T ACGTG
TTGTACGTG
TTGCACGTG
18. Once paired with a complement after strand invasion, the end, unlike a end, can be extended by a DNA polymerase.
19. During homologous genetic recombination, a Holliday intermediate may be formed almost anywhere within the two paired, homologous chromosomes; the branch point of the intermediate can move extensively by branch migration. In site-specific recombination, the Holliday intermediate is formed between two specific sites, and branch migration is generally restricted by heterologous sequences on either side of the recombination sites.
20.
Points Y
Points X
21. Once replication has proceeded from the origin to a point where one recombination site has been replicated but the other has not, site-specific recombination not only inverts the DNA between the recombination sites but also changes the direction of one replication fork relative to the other. The forks will chase each other around the DNA circle, generating many tandem copies of the plasmid. The multimeric circle can be resolved to monomers by additional site-specific recombination events.
22.
Even in the absence of an added mutagen, background mutations occur due to radiation, cellular chemical reactions, and so forth.
If the DNA is sufficiently damaged, a substantial fraction of gene products is nonfunctional and the cell is nonviable.
Cells with reduced DNA repair capability are more sensitive to mutagens. Because they less readily repair lesions caused by R7000, bacteria have an increased mutation rate and increased chance of lethal effects.
In the strain, the excision-repair system removes DNA bases with attached , decreasing the amount of in these cells over time. In the , the DNA is not repaired and the level increases as continues to react with the DNA.
All mutations listed in the table except to show significant increases over background. Each type of mutation results from a different type of interaction between R7000 and DNA. Because different types of interactions are not equally likely (due to differences in reactivity, steric constraints, etc.), the resulting mutations occur with different frequencies.
No. Only those that start with a base pair are explained by this model. Thus to and to must be due to R7000 attaching to an A or a T.
R7000―G pairs with A. First, R7000 adds to to give . (Compare this with what happens with the in Fig. 25-27b.) If this is not repaired, one strand is replicated as , which is repaired to . The other strand is wild-type. If the replication produces , a similar pathway leads to an base pair.
No. Compare data in the two tables, and keep in mind that different mutations occur at different frequencies.
to : moderate in both strains; but better repair in
to : moderate in both; no real difference
to : higher in ; certainly less repair!
to : high in both; no real difference
to : high in both; no real difference
to : low in both; no real difference
Certain adducts may be more readily recognized by the repair apparatus than others, and these are repaired more rapidly and result in fewer mutations.
Chapter 26
1.
60 to 100 s
500 to 900 nucleotides
6 to 11 h
2. A single base error in DNA replication, if not corrected, would cause one of the two daughter cells, and all its progeny, to have a mutated chromosome. A single base error in RNA transcription would not affect the chromosome; it would lead to formation of some defective copies of one protein, but because mRNAs turn over rapidly, most copies of the protein would not be defective. The progeny of this cell would be normal.
3. Normal posttranscriptional processing at the end (cleavage and polyadenylation) would be inhibited or blocked.
4. Because the template-strand RNA does not encode the enzymes needed to initiate viral infection, it would probably be inert or simply degraded by cellular ribonucleases. Replication of the template-strand RNA and propagation of the virus could occur only if intact RNA replicase (RNA-dependent RNA polymerase) were introduced into the cell along with the template strand.
5. AUGUCCAAAAUCGUA
6. (1) Use of a template strand of nucleic acid; (2) synthesis in the direction; (3) use of nucleoside triphosphate substrates, with formation of a phosphodiester bond and displacement of . Polynucleotide phosphorylase forms phosphodiester bonds but differs in all other listed properties.
7. Most of the RNA transcribed in the nucleus was intronic and removed from the mRNAs.
8. No, at this time it is not possible to determine a cell’s transcriptome based solely on the genome. Different cells have different transcriptomes based on which promoters are being used and how the transcripts are processed by factors present in the cell.
9. Generally two: one to cleave the phosphodiester bond at one intron-exon junction, the other to link the resulting free exon end to the exon at the other end of the intron. If the nucleophile in the first step were water, this step would be a hydrolysis, and only one transesterification step would be required to complete the splicing process.
10. Many snoRNAs, required for rRNA modification reactions, are encoded in introns. If splicing does not occur, snoRNAs are not produced.
11.
Water attacks the C-6 position of adenine, forming a tetrahedral intermediate, which then eliminates ammonia to form inosine.
Inosine can no longer pair correctly with U residues found on the opposite RNA strand prior to ADAR activity. This results in disruption of the RNA duplex.
Inosine does not have the same pairing properties as adenine and could potentially recode that particular codon so that a different amino acid is incorporated into the protein.
12. Physical separation of the processes prevents translation of primary or precursor transcripts that have not yet been processed by the cell. It also prevents the RNA processing and translational machineries from competing with one another for mRNAs.
13.
This could change how proteins or other RNAs recognize a particular RNA sequence and interfere with splicing, poly(A) formation, RNA modification, or other steps in which sequence-specific RNA recognition is important.
S-adenosylmethionine serves as the methyl donor in the synthesis of -methyladenosine.
14. These enzymes lack a proofreading exonuclease and have a high error rate; the likelihood of a replication error that would inactivate the virus is much lower in a small genome than in a large one.
15.
0.00002%
For the “unnatural selection” step, use a chromatographic resin with a bound molecule that is a transition-state analog of the ester hydrolysis reaction.
16. Though RNA synthesis is quickly halted by -amanitin toxin, it takes several days for the critical mRNAs and proteins in the liver to degrade, causing liver dysfunction and death.
17.
After lysis of the cells and partial purification of the contents, an antibody-based assay could detect the subunit. The subunit could then be subjected to tandem mass spectrometry, which could detect the difference in amino acid residues between the normal subunit and the mutated form.
Direct DNA sequencing (by the Sanger method)
18.
384
1,620 nucleotide pairs
Most of the nucleotides are untranslated regions at the and ends of the mRNA. Also, most mRNAs code for a signal sequence (Chapter 27) in their protein products, which is eventually removed to produce the mature, functional protein.
19.
Injection of the antisense oligo cleaves the c-mos RNA and removes its poly(A) tail. This correlates with a loss in oocyte maturation. The noncoding poly(A) tail of the c-mos mRNA must be important for its function in maturation.
The sense oligo control shows that these results depend on complementarity to the c-mos mRNA.
The prosthetic RNA likely base-paired with the amputated c-mos mRNA and restored its function. Therefore, poly(A) tails can function “in trans” — they do not need to be covalently connected to the mRNAs they regulate.
The poly(A) tail stimulates protein expression in the reporter. This is likely also occurring with c-mos, and this protein expression is important for oocyte maturation.
These results suggest that the expression of genes can be artificially controlled either by cutting off their poly(A) tails or by attaching synthetic ones derived from other genes. This could be useful for fine-tuning gene expression in bioengineered cells or turning expression on or off.
Chapter 27
1.
in eukaryotes; in bacteria
2. UUAAUGUAU, UUGAUGUAU, CUUAUGUAU, CUCAUGUAU, CUAAUGUAU, CUGAUGUAU, UUAAUGUAC, UUGAUGUAC, CUUAUGUAC, CUCAUGUAC, CUAAUGUAC, CUGAUGUAC
3. No. Because nearly all the amino acids have more than one codon (e.g., Leu has six), any given polypeptide can be encoded by several different base sequences. However, some amino acids are encoded by only one codon, and those with multiple codons often share the same nucleotide at two of the three positions, so certain parts of the mRNA sequence encoding a protein of known amino acid sequence can be predicted with high certainty.
4.
No. The complementary antiparallel strands in double-helical DNA do not have the same base sequence in the direction. RNA is transcribed from only one specific strand of duplex DNA. The RNA polymerase must therefore recognize and bind to the correct strand.
5. (a), (b), and (c) are correct.
This outcome would restore the original gene and allow production of the native protein.
Altering the gene to insert a different amino acid at this position would allow for the generation of a full-length protein. Some activity may be present, especially if the new amino acid represented a conservative alteration (such as Ile substituting for Val).
This outcome would be similar to (c), inserting an amino acid (probably a different one) at the affected position and allowing synthesis of a full length and perhaps active protein. This outcome is called nonsense suppression. It works because most cells have multiple copies of particular tRNAs, some of which are expressed at low levels. If a minor one is altered, the genetic code is not disrupted because the other copies of the tRNA provide normal function.
This outcome would rarely work, since it would tend to introduce too many amino acid changes into the protein.
6. The labeled amino acids were found at the carboxyl end. Dintzis only isolated complete α subunits. With short incubation times, labeled amino acids would only appear in the part of the polypeptide that was synthesized last. Labeled amino acids introduced at the amino terminus would not be seen, because those polypeptides would not have been completed prior to protein isolation.
7. There are two tRNAs for methionine: , which is the initiating tRNA, and , which can insert a Met residue in interior positions in a polypeptide. Only is recognized by the initiation factor IF2 and is aligned with the initiating AUG positioned at the ribosomal P site in the initiation complex. AUG codons in the interior of the mRNA can bind and incorporate only .
8. is translated as . The peptide is 14 amino acids long instead of 15, because the last codon is a stop codon. Ten tRNAs are needed, one for each type of amino acid.
9. Allow polynucleotide phosphorylase to act on a mixture of UDP and CDP in which UDP has, say, five times the concentration of CDP. The result would be a synthetic RNA polymer with many UUU triplets (coding for Phe), a smaller number of UUC (Phe), UCU (Ser), and CUU (Leu), a much smaller number of UCC (Ser), CUC (Leu), and CCU (Pro), and an even smaller number of CCC (Pro).
10. A minimum of 583 ATP equivalents (based on 4 per amino acid residue added, except that there are only 145 translocation steps). Correction of each error requires 2 ATP equivalents. For glycogen synthesis, 292 ATP equivalents are required. The extra energy cost for -globin synthesis reflects the cost of the information content of the protein. At least 20 activating enzymes, 70 ribosomal proteins, 4 rRNAs, 32 or more tRNAs, an mRNA, and 10 or more auxiliary enzymes must be made by the eukaryotic cell in order to synthesize a protein from amino acids. The synthesis of an chain of glycogen from glucose requires only 4 or 5 enzymes (Chapter 15).
11.
Glycine codons Anticodons The and middle position
Pairings with anticodons , and UCC
Pairings with anticodons and CCC
12. (a), (c), (e), and (g) only; (b), (d), and (f) cannot be the result of single-base mutations, because (b) and (f) would require substitutions of two bases, and (d) would require substitutions of all three bases.
13.
Changes: CC AU U C C U C A A G G G Of 63 possible one-base changes, 14 would result in no coding change.
14. The two DNA codons for Glu are GAA and GAG, and the four DNA codons for Val are GTT, GTC, GTA, and GTG. A single base change in GAA to form GTA or in GAG to form GTG could account for the replacement in sickle-cell hemoglobin. Much less likely are two-base changes, from GAA to GTG, GTT, or GTC; and from GAG to GTA, GTT, or GTC.
15. Isoleucine is similar in structure to several other amino acids, particularly valine. Distinguishing between valine and isoleucine in the aminoacylation process requires a second filter — a proofreading function. Histidine has a structure unlike that of any other amino acid, providing opportunities for binding specificity adequate to ensure accurate aminoacylation of the cognate tRNA.
16.
The Ala-tRNA synthetase recognizes the base pair in the amino acid arm of .
The mutant would insert Ala residues at codons encoding Pro.
A mutation that might have similar effects is an alteration in that allowed it to be recognized and aminoacylated by Ala-tRNA synthetase.
Most of the proteins in the cell would be inactivated, so these would be lethal mutations and hence never observed. This represents a powerful selective pressure for maintaining the genetic code.
17. The 15,000 ribosomes in an E. coli cell can synthesize more than 23,000 proteins in 20 minutes.
18. IF2: The 70S ribosome would form, but initiation factors would not be released and elongation could not start. EF-Tu: The second aminoacyl-tRNA would bind to the ribosomal A site, but no peptide bond would form. EF-G: The first peptide bond would form, but the ribosome would not move along the mRNA to vacate the A site for binding of a new EF-Tu–tRNA.
19. The amino acid most recently added to a growing polypeptide chain is the only one covalently attached to a tRNA and thus is the only link between the polypeptide and the mRNA encoding it. A proofreading activity would sever this link, halting synthesis of the polypeptide and releasing it from the mRNA.
20. SecA; Drugs that inhibit the ability of SecA to bind bacterial proteins or hydrolyze ATP could significantly disrupt the export of bacterial proteins. SecB is nonessential as bacteria have numerous other chaperone proteins. Antibiotics that target the SecYEG complex, which is homologous to its eukaryotic counterpart (Sec61), could cause significant adverse effects in humans.
21. The protein would be directed into the ER, and from there the targeting would depend on additional signals. SRP binds the amino-terminal signal early in protein synthesis and directs the nascent polypeptide and ribosome to receptors in the ER. Because the protein is translocated into the lumen of the ER as it is synthesized, the NLS is never accessible to the proteins involved in nuclear targeting.
22. Trigger factor is a molecular chaperone that stabilizes an unfolded and translocation-competent conformation of ProOmpA.
23. DNA with a minimum of 5,784 bp; some of the coding sequences must be nested or overlapping.
24.
The helices associate through the hydrophobic effect and van der Waals interactions.
R groups 3, 6, 7, and 10 extend to the left; 1, 2, 4, 5, 8, and 9 extend to the right.
One possible sequence is
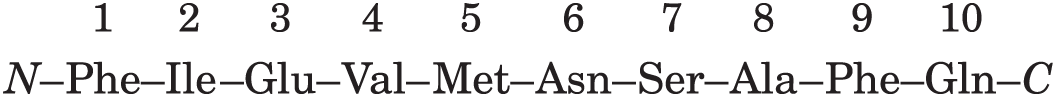
One possible DNA sequence for the amino acid sequence in (c) is
Phe, Leu, Ile, Met, and Val. All are hydrophobic, but the set does not include all the hydrophobic amino acids; Trp, Pro, and Ala are missing.
Tyr, His, Gln, Asn, Lys, Asp, and Glu. All of these are hydrophilic, although Tyr is less hydrophilic than the others. The set does not include all the hydrophilic amino acids; Ser, Thr, and Arg are missing.
Omitting T from the mixture excludes codons starting or ending with T — thus excluding Tyr, which is not very hydrophilic, and, more importantly, excluding the two possible stop codons (TAA and TAG). No other amino acids in the NAN set are excluded by omitting T.
Misfolded proteins are often degraded in the cell. Therefore, if a synthetic gene has produced a protein that forms a band on the SDS gel, it is likely that this protein is folded properly.
Protein folding depends on more than the hydrophobic effect and van der Waals interactions. There are many reasons why a synthesized random-sequence protein might not fold into the four-helix structure. For example, hydrogen bonds between hydrophilic side chains could disrupt the structure. Also, not all sequences have an equal propensity to form an α helix.
Chapter 28
1.
Tryptophan synthase levels remain high despite the presence of tryptophan.
Levels again remain high.
Levels rapidly decrease, preventing wasteful synthesis of tryptophan.
2. The E. coli cells will produce -galactosidase when they are subjected to high levels of a DNA-damaging agent such as UV light. Under such conditions, RecA binds to single-stranded chromosomal DNA and facilitates autocatalytic cleavage of the LexA repressor, releasing LexA from its binding site and allowing transcription of downstream genes.
3.
Constitutive, low-level expression of the operon; most mutations in the operator would make the repressor less likely to bind.
Constitutive expression; mutation prevents negative regulation of the operon.
Increased expression; under conditions in which the operon is induced, mutation increases recruitment of RNA polymerase.
Constant repression; mutation allows repressor to readily bind to operator.
Decreased expression; under conditions in which the operon is induced, mutation decreases recruitment of RNA polymerase.
4. 7,000 copies
5. , about times greater than the dissociation constant. With 10 copies of active repressor in the cell, the operator site is always bound by the repressor molecule.
6. (a) through (e). Each condition decreases expression of lac operon genes.
7.
Less attenuation. The ribosome completing the translation of sequence 1 would no longer overlap and block sequence 2; sequence 2 would always be available to pair with sequence 3, preventing formation of the attenuator structure.
More attenuation. Sequence 2 would pair less efficiently with sequence 3; the attenuator structure would be formed more often, even when sequence 2 was not blocked by a ribosome.
No attenuation. The only regulation would be that afforded by the Trp repressor.
Attenuation loses its sensitivity to Trp tRNA. It might become sensitive to His tRNA.
Attenuation would rarely, if ever, occur. Sequences 2 and 3 always block formation of the attenuator.
Constant attenuation. Attenuator always forms, regardless of the availability of tryptophan.
8. Induction of the SOS response could not occur, making the cells more sensitive to high levels of DNA damage.
9. Each Salmonella cell would have flagella made up of both types of flagellar protein, and the cell would be vulnerable to antibodies generated in response to either protein.
10. A dissociable factor necessary for activity (e.g., a specificity factor similar to the subunit of the E. coli enzyme) may have been lost during purification of the polymerase.
11.
Gal4 protein
Gal4p DNA-binding domain
Gal4p activator domain
Engineered protein
Lac repressor DNA-binding domain
Gal4p activator domain
The engineered protein cannot bind to the Gal4p-binding site in the GAL gene because it lacks the Gal4p DNA-binding domain. Modify the Gal4p-binding site in the DNA to give it the nucleotide sequence to which the Lac repressor normally binds (using methods described in Chapter 9).
12. Methylamine. The reaction proceeds with attack of water on the guanidinium carbon of the modified arginine.
13. Synthesis of a protein first requires the synthesis of an mRNA long enough to encode the protein and to include any necessary regulatory sequences, with one ribonucleoside triphosphate used up for every nucleotide residue included in the mRNA. Then, the mRNA must be translated — one of the most energy-intensive processes in the cell (Chapter 27). To maintain repression, the repressor protein would need to be synthesized repeatedly. With the use of RNA as repressor, the RNA can be shorter than a protein repressor–encoding mRNA, and no translation step is required.
14. The bcd mRNA needed for development is contributed to the egg by the mother. The fertilized egg develops normally even if its genotype is , as long as the mother has one normal bcd gene and the allele is recessive. However, the adult female will be sterile because she has no normal bcd mRNA to contribute to her eggs.
15.
For 10% expression (90% repression), 10% of the repressor has bound inducer and 90% is free and available to bind the operator. The calculation uses Eqn 5-8 (p. 151), with and :
For 90% expression, 90% of the repressor has bound inducer, so . Entering the values for Y and in Eqn 5-8 gives . Thus, gene expression varies 10-fold over a roughly 10-fold [IPTG] range.
You would expect the protein levels to be low before induction, to rise during induction, and then to decay as synthesis stops and the proteins are degraded.
As shown in (a), the lac operon has more levels of expression than just on or off; thus it does not have characteristic A. As shown in (b), expression of the lac operon subsides once the inducer is removed; thus it lacks characteristic B.
GFP-on: (designating the protein product of the gene) and GFP are expressed at high levels; represses , so no LacI is produced. GFP-off: LacI is expressed at a high level; LacI represses , so and GFP are not produced.
IPTG treatment switches the system from GFP-off to GFP-on. IPTG has an effect only when LacI is present, so it affects only the GFP-off state. Adding IPTG relieves the repression of , allowing high-level expression of , which turns off expression of LacI, and high-level expression of GFP.
Heat treatment switches the system from GFP-on to GFP-off. Heat has an effect only when is present, so affects only the GFP-on state. Heat inactivates and relieves the repression of , allowing high-level expression of LacI. LacI then acts at to repress synthesis of and GFP.
Characteristic A: The system is not stable in the intermediate state. At some point, one repressor will act more strongly than the other due to chance fluctuations in expression; this shuts off expression of the other repressor and locks the system in one state. Characteristic B: Once one repressor is expressed, it prevents the synthesis of the other; thus the system remains in one state even after the switching stimulus has been removed.
At no time does any cell express an intermediate level of GFP—this is a confirmation of characteristic A. At the intermediate concentration (X) of inducer, some cells have switched to GFP-on while others have not yet made the switch and remain in the GFP-off state; none are in between. The bimodal distribution of expression levels at is caused by the mixed population of GFP-on and GFP-off cells.