Chapter Review
Key Terms
Terms in bold are defined in the glossary.
- cellular respiration
- citric acid cycle
- tricarboxylic acid (TCA) cycle
- Krebs cycle
- thioester
- mitchondrial pyruvate carrier (MPC)
- pyruvate dehydrogenase (PDH) complex
- oxidative decarboxylation
- lipoate
- substrate channeling
- iron-sulfur center
- α-ketoglutarate dehydrogenase complex
- moonlighting enzymes
- nucleoside diphosphate kinase
- prochiral molecule
- amphibolic pathway
- glyoxylate cycle
- anaplerotic reaction
- biotin
- oncometabolite
- metabolon
Problems
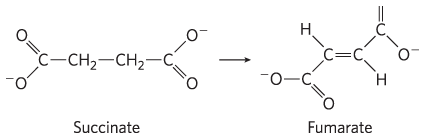
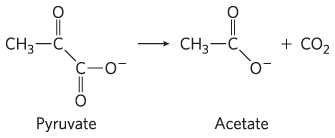
1. Balance Sheet for the Citric Acid Cycle The citric acid cycle has eight enzymes: citrate synthase, aconitase, isocitrate dehydrogenase, α-ketoglutarate dehydrogenase, succinyl-CoA synthetase, succinate dehydrogenase, fumarase, and malate dehydrogenase.
- Write a balanced equation for the reaction catalyzed by each enzyme.
- Name the cofactor(s) required by each enzyme reaction.
- For each enzyme, determine which of the following describes the type of reaction(s) catalyzed: condensation (carbon–carbon bond formation); dehydration (loss of water); hydration (addition of water); decarboxylation (loss of ); oxidation-reduction; substrate-level phosphorylation; isomerization.
- Write a balanced net equation for the catabolism of acetyl-CoA to .
2. Net Equation for Glycolysis and the Citric Acid Cycle Write the net biochemical equation for the metabolism of a molecule of glucose by glycolysis and the citric acid cycle, including all cofactors.
3. Recognizing Oxidation and Reduction Reactions One biochemical strategy of many living organisms is the stepwise oxidation of organic compounds to and and the conservation of a major part of the energy thus produced in the form of ATP. It is important to be able to recognize oxidation-reduction processes in metabolism. Reduction of an organic molecule results from the hydrogenation of a double bond (Eqn 1, below) or of a single bond with accompanying cleavage (Eqn 2). Conversely, oxidation results from dehydrogenation. In biochemical redox reactions, the coenzymes NAD and FAD organic molecules in the presence of the proper enzymes.
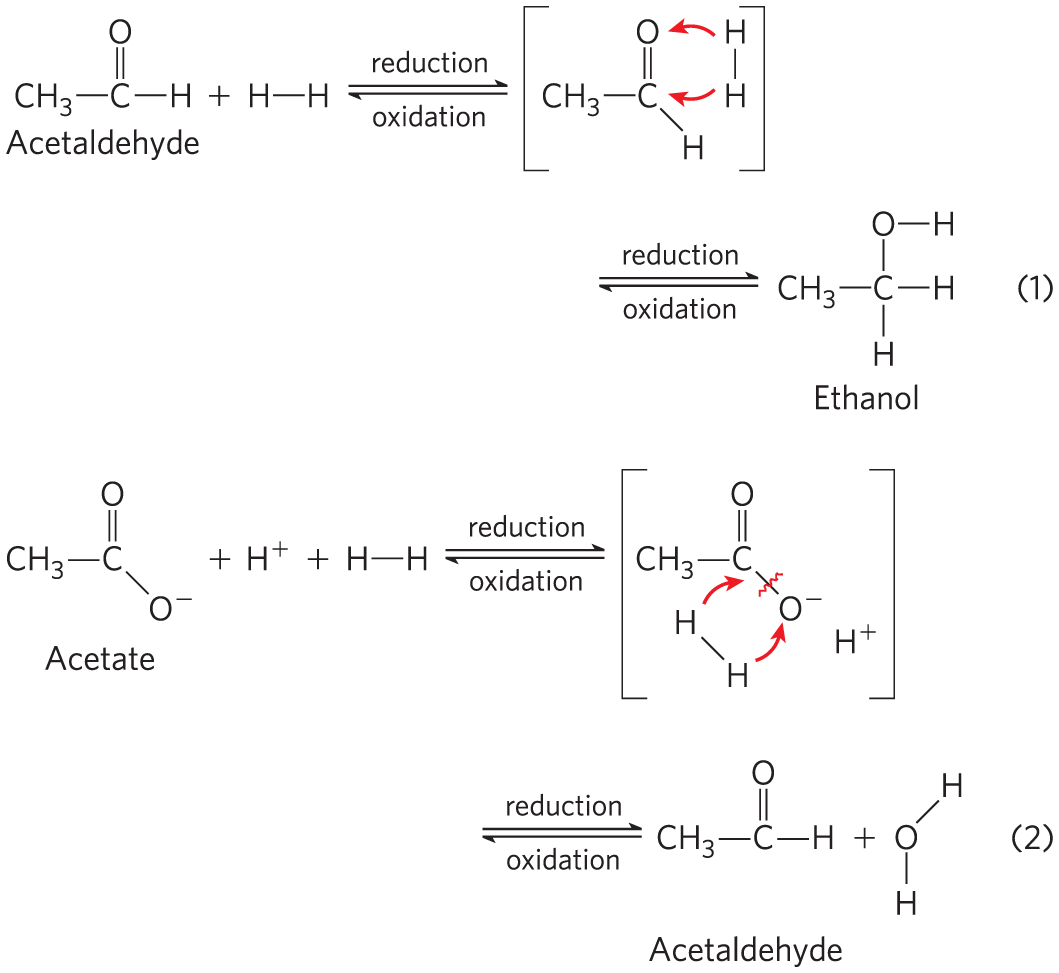
For each of the metabolic transformations (a) through (h), determine whether the compound on the left has undergone oxidation or reduction. Balance each transformation by inserting and, where necessary, .
4. Relationship between Energy Release and the Oxidation State of Carbon A eukaryotic cell can use glucose and hexanoate as fuels for cellular respiration. On the basis of their structural formulas, which substance releases more energy per gram on complete combustion to and ?
5. Nicotinamide Coenzymes as Reversible Redox Carriers The nicotinamide coenzymes (see Fig. 13-24) can undergo reversible oxidation-reduction reactions with specific substrates in the presence of the appropriate dehydrogenase. In these reactions, serves as the hydrogen source, as described in Problem 3. Whenever the coenzyme is oxidized, a substrate must be simultaneously reduced:
For each of the reactions in (a) through (f) shown below, determine whether the substrate has been oxidized or reduced or is unchanged in oxidation state (see Problem 3). If a redox change has occurred, balance the reaction with the necessary amount of , NADH, , and . The objective is to recognize when a redox coenzyme is necessary in a metabolic reaction.
6. Pyruvate Dehydrogenase Cofactors and Mechanism Describe the role of each cofactor involved in the reaction catalyzed by the pyruvate dehydrogenase complex.
7. Thiamine Deficiency Individuals with a thiamine-deficient diet have relatively high levels of pyruvate in their blood. Explain this in biochemical terms.
8. Isocitrate Dehydrogenase Reaction What type of chemical reaction is involved in the conversion of isocitrate to α-ketoglutarate? Name and describe the role of any cofactors. What other reaction(s) of the citric acid cycle are of this same type?
9. Stimulation of Oxygen Consumption by Oxaloacetate and Malate In the early 1930s, Albert Szent-Györgyi reported the interesting observation that the addition of small amounts of oxaloacetate or malate to suspensions of minced pigeon breast muscle stimulated the oxygen consumption of the preparation. Surprisingly, the amount of oxygen consumed was about seven times more than the amount necessary for complete oxidation (to and ) of the added oxaloacetate or malate. Why did the addition of oxaloacetate or malate stimulate oxygen consumption? Why was the amount of oxygen consumed so much greater than the amount necessary to completely oxidize the added oxaloacetate or malate?
10. Formation of Oxaloacetate in a Mitochondrion In the last reaction of the citric acid cycle, malate is dehydrogenated to regenerate the oxaloacetate necessary for the entry of acetyl-CoA into the cycle:
- Calculate the equilibrium constant for this reaction at 25 °C.
- Because assumes a standard pH of 7, the equilibrium constant calculated in (a) corresponds to
The measured concentration of l-malate in rat liver mitochondria is about 0.20 mm when is 10. Calculate the concentration of oxaloacetate at pH 7 in these mitochondria.
- To appreciate the magnitude of the mitochondrial oxaloacetate concentration, calculate the number of oxaloacetate molecules in a single rat liver mitochondrion. Assume the mitochondrion is a sphere of diameter .
11. Cofactors for the Citric Acid Cycle Suppose you have prepared a mitochondrial extract that contains all the soluble enzymes of the matrix but has lost (by dialysis) all the low molecular weight cofactors. What must you add to the extract so that the preparation will oxidize acetyl-CoA to ?
12. Riboflavin Deficiency How would a riboflavin deficiency affect the functioning of the citric acid cycle? Explain your answer.
13. Oxaloacetate Pool What factors might decrease the pool of oxaloacetate available for the activity of the citric acid cycle? How can the pool of oxaloacetate be replenished?
14. Energy Yield from the Citric Acid Cycle The reaction catalyzed by succinyl-CoA synthetase produces the high-energy compound GTP. How is the free energy contained in GTP incorporated into the cellular ATP pool?
15. Respiration Studies in Isolated Mitochondria Cellular respiration can be studied in isolated mitochondria by measuring oxygen consumption under different conditions. If 0.01 m sodium malonate is added to actively respiring mitochondria that are using pyruvate as fuel, respiration soon stops and a metabolic intermediate accumulates.
- What is the structure of this intermediate?
- Explain why it accumulates.
- Explain why oxygen consumption stops.
- Aside from removal of the malonate, what can overcome this inhibition of respiration? Explain.
16. Labeling Studies in Isolated Mitochondria Biochemists have often delineated the metabolic pathways of organic compounds by using a radioactively labeled substrate and following the fate of the label.
- How can you determine whether a suspension of isolated mitochondria metabolizes added glucose to and ?
- Suppose you add a brief pulse of (labeled in the methyl position) to the mitochondria. After one turn of the citric acid cycle, what is the location of the in the oxaloacetate? Explain by tracing the label through the pathway. How many turns of the cycle are required to release all the as ?
17. An investigator briefly incubates an actively respiring bacterial culture with glucose and isolates the glycolytic and citric acid cycle intermediates. Where is the located in each of the intermediates listed? Consider only the initial incorporation of in the first pass of labeled glucose through the pathways.
- Fructose 1,6-bisphosphate
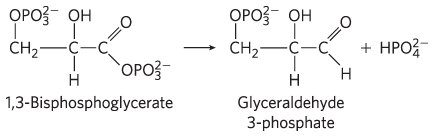
- Glyceraldehyde 3-phosphate
- Phosphoenolpyruvate
- Acetyl-CoA
- Citrate
- α-Ketoglutarate
- Oxaloacetate
18. Role of the Vitamin Thiamine People with beriberi, a disease caused by thiamine deficiency, have elevated levels of blood pyruvate and α-ketoglutarate, especially after consuming a meal rich in glucose. How are these effects related to a deficiency of thiamine?
19. Synthesis of Oxaloacetate by the Citric Acid Cycle In the last step of the citric acid cycle, -dependent oxidation of l-malate forms oxaloacetate. Can a net synthesis of oxaloacetate from acetyl-CoA occur using only the enzymes and cofactors of the citric acid cycle, without depleting the intermediates of the cycle? Explain. How do cells replenish the oxaloacetate that is lost from the cycle to biosynthetic reactions?
20. Oxaloacetate Depletion Mammalian liver can carry out gluconeogenesis using oxaloacetate as the starting material (Chapter 14). Would the extensive use of oxaloacetate for gluconeogenesis affect the operation of the citric acid cycle? Explain your answer.
21. Mode of Action of the Rodenticide Fluoroacetate Fluoroacetate, prepared commercially for rodent control, is also produced by a South African plant. After entering a cell, fluoroacetate is converted to fluoroacetyl-CoA in a reaction catalyzed by the enzyme acetate thiokinase:
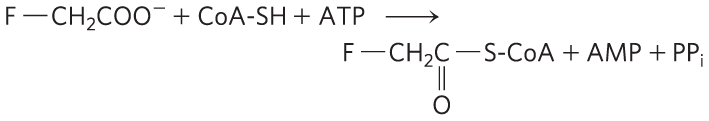
You perform a perfusion experiment to study the toxic effect of fluoroacetate using intact isolated rat heart. After perfusing the heart with 0.22 mm fluoroacetate, you see a decrease in the measured rate of glucose uptake and glycolysis as well as an accumulation of glucose 6-phosphate and fructose 6-phosphate. Examination of the citric acid cycle intermediates reveals that their concentrations are below normal, except for citrate, which has a concentration 10 times higher than normal.
- Where did the block in the citric acid cycle occur? What caused citrate to accumulate and the other cycle intermediates to be depleted?
- Fluoroacetyl-CoA is enzymatically transformed in the citric acid cycle. What is the structure of the end product of fluoroacetate metabolism? Why does it block the citric acid cycle? How might the inhibition be overcome?
- In the heart perfusion experiments, why did glucose uptake and glycolysis decrease? Why did hexose monophosphates accumulate?
- Why is fluoroacetate poisoning fatal?
22. Synthesis of l-Malate in Wine Making The tartness of some wines is due to high concentrations of l-malate. Write a sequence of reactions showing how yeast cells synthesize l-malate from glucose under anaerobic conditions in the presence of dissolved Note that the overall reaction for this fermentation cannot involve the consumption of nicotinamide coenzymes or citric acid cycle intermediates.
23. Net Synthesis of α-Ketoglutarate α-Ketoglutarate plays a central role in the biosynthesis of several amino acids. Write a sequence of enzymatic reactions that could result in the net synthesis of α-ketoglutarate from pyruvate. Your proposed sequence must not involve the net consumption of other citric acid cycle intermediates. Write an equation for the overall reaction.
24. Amphibolic Pathways Explain, giving examples, what is meant by the statement that the citric acid cycle is amphibolic.
25. Regulation of the Pyruvate Dehydrogenase Complex In animal tissues, the ratio of active, unphosphorylated to inactive, phosphorylated PDH complex regulates the rate of conversion of pyruvate to acetyl-CoA. Determine what happens to the rate of this reaction when a preparation of rabbit muscle mitochondria containing the PDH complex is treated with (a) pyruvate dehydrogenase kinase, ATP, and NADH; (b) pyruvate dehydrogenase phosphatase and ; (c) malonate.
26. Commercial Synthesis of Citric Acid Manufacturers use citric acid as a flavoring agent in soft drinks, fruit juices, and many other foods. Worldwide, the market for citric acid is valued at hundreds of millions of dollars per year. Commercial production uses the mold Aspergillus niger, which metabolizes sucrose under carefully controlled conditions.
- The yield of citric acid strongly depends on the concentration of in the culture medium, as indicated in the graph. Why does the yield decrease when the concentration of is above or below the optimal value of 0.5 mg/L?
![A graph plots a curve showing yield of citric acid produced by a mold against the concentration of [F e C l 3] in the growth medium.](../images/8e_16_un16.png)
- Write the sequence of reactions by which A. niger synthesizes citric acid from sucrose. Write an equation for the overall reaction.
- Does the commercial process require the culture medium to be aerated — that is, is this a fermentation or an aerobic process? Explain.
- The yield of citric acid strongly depends on the concentration of in the culture medium, as indicated in the graph. Why does the yield decrease when the concentration of is above or below the optimal value of 0.5 mg/L?
27. Regulation of Citrate Synthase In the presence of saturating amounts of oxaloacetate, the activity of citrate synthase from pig heart tissue shows a sigmoid dependence on the concentration of acetyl-CoA, as shown in the graph. Adding succinyl-CoA shifts the curve to the right and makes the sigmoid dependence more pronounced.
![A graph plots [Acetyl-Co A] against activity and shows two curves, one for no succinyl-Co A and one for succinyl-Co A added.](../images/8e_16_un17.png)
On the basis of these observations, suggest how succinyl-CoA regulates the activity of citrate synthase. (Hint: See Fig. 6-37.) Why is succinyl-CoA an appropriate signal for regulation of the citric acid cycle? How does the regulation of citrate synthase control the rate of cellular respiration in pig heart tissue?
28. Regulation of Pyruvate Carboxylase The carboxylation of pyruvate by pyruvate carboxylase occurs at a very low rate unless acetyl-CoA, a positive allosteric modulator, is present. If you have just eaten a meal rich in fatty acids (triacylglycerols) but low in carbohydrates (glucose), how does this regulatory property shut down the oxidation of glucose to and but increase the oxidation of acetyl-CoA derived from fatty acids?
29. Relationship between Respiration and the Citric Acid Cycle Although oxygen does not participate directly in the citric acid cycle, the cycle operates only when is present. Why?
30. Effect of on the Citric Acid Cycle How would you expect the operation of the citric acid cycle to respond to a rapid increase in the ratio in the mitochondrial matrix? Why?
31. Thermodynamics of Citrate Synthase Reaction in Cells Citrate is formed by the condensation of acetyl-CoA with oxaloacetate, catalyzed by citrate synthase:
In rat heart mitochondria at pH 7.0 and 25 °C, the concentrations of reactants and products are oxaloacetate, ; acetyl-CoA, ; citrate, ; and CoA, . The standard free-energy change for the citrate synthase reaction is . What is the direction of metabolite flow through the citrate synthase reaction in rat heart cells? Explain.
32. Reactions of the Pyruvate Dehydrogenase Complex Two of the steps in the oxidative decarboxylation of pyruvate (steps and in Fig. 16-6) do not involve any of the three carbons of pyruvate, yet are essential to the operation of the PDH complex. Explain.
33. Pyruvate Transport into Mitochondria The mitochondrial pyruvate carrier (MPC) is a heterodimer of the proteins MPC1 and MPC2. In a high proportion (80%) of certain cancers, including gliomas (tumors of the glial cells of the brain), the gene for one of these proteins is mutated such that pyruvate cannot enter the mitochondrial matrix. Name three metabolic effects that you would expect to see if cytosolic pyruvate could not gain access to the machinery of the citric acid cycle. (Hint: Box 14-1 may be helpful.)
34. Citric Acid Cycle Mutants There are many cases of human disease in which one or another enzyme activity is lacking due to genetic mutation. Why are cases in which individuals lack one of the enzymes of the citric acid cycle extremely rare?
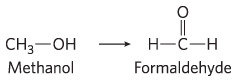
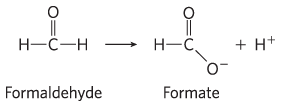
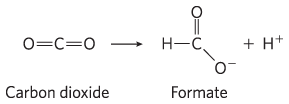
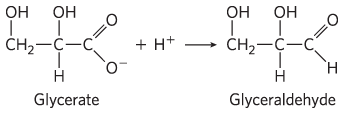
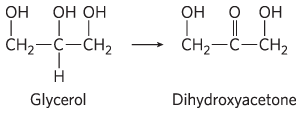
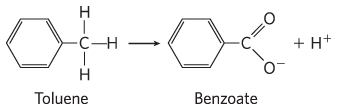
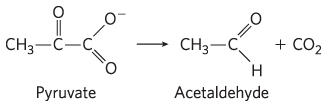
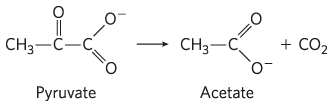
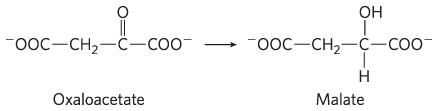
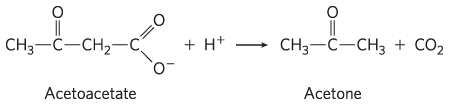

 and
and  in
in Data Analysis Problem
35. How the Citric Acid Cycle Was Discovered The detailed biochemistry of the citric acid cycle was determined by several researchers over a period of decades. In a 1937 article, Krebs and Johnson summarized their work and the work of others in the first published description of this pathway.
The methods used by these researchers were very different from those of modern biochemistry. Radioactive tracers were not commonly available until the 1940s, so Krebs and other researchers had to use nontracer techniques to work out the pathway. Using freshly prepared samples of pigeon breast muscle, they determined oxygen consumption by suspending minced muscle in buffer in a sealed flask and measuring the volume (in μL) of oxygen consumed under different conditions. They measured levels of substrates (intermediates) by treating samples with acid to remove contaminating proteins, then assaying the quantities of various small organic molecules. The two key observations that led Krebs and colleagues to propose a citric acid cycle as opposed to a linear pathway (like that of glycolysis) were made in the following experiments.
Experiment I: They incubated 460 mg of minced muscle in 3 mL of buffer at 40 °C for 150 minutes. Addition of citrate increased consumption by 893 μL compared with samples without added citrate. They calculated, based on the consumed during respiration of other carbon-containing compounds, that the expected consumption for complete respiration of this quantity of citrate was only 302 μL.
Experiment II: They measured consumption by 460 mg of minced muscle in 3 mL of buffer when incubated with citrate and/or with 1-phosphoglycerol (glycerol 1-phosphate; this was known to be readily oxidized by cellular respiration) at 40 °C for 140 minutes. The results are shown in the table.
Sample Substrate(s) added absorbed 1
No extra
342
2
0.3 mL 0.2 m 1-phosphoglycerol
757
3
0.15 mL 0.02 m citrate
431
4
0.3 mL 0.2 m 1-phosphoglycerol and 0.15 mL 0.02 m citrate
1,385
- Why is consumption a good measure of cellular respiration?
- Why does sample 1 (unsupplemented muscle tissue) consume some oxygen?
- Based on the results for samples 2 and 3, can you conclude that 1-phosphoglycerol and citrate serve as substrates for cellular respiration in this system? Explain your reasoning.
- Krebs and colleagues used the results from these experiments to argue that citrate was “catalytic” — that it helped the muscle tissue samples metabolize 1-phosphoglycerol more completely. How would you use their data to make this argument?
- Krebs and colleagues further argued that citrate was not simply consumed by these reactions, but had to be regenerated. Therefore, the reactions had to be a cycle rather than a linear pathway. How would you make this argument?
Other researchers had found that arsenate inhibits α-ketoglutarate dehydrogenase and that malonate inhibits succinate dehydrogenase.
- Krebs and coworkers found that muscle tissue samples treated with arsenate and citrate would consume citrate only in the presence of oxygen; under these conditions, oxygen was consumed. Based on the pathway in Figure 16-7, what was the citrate converted to in this experiment, and why did the samples consume oxygen?
In their article, Krebs and Johnson further reported the following: (1) In the presence of arsenate, 5.48 mmol of citrate was converted to 5.07 mmol of α-ketoglutarate. (2) In the presence of malonate, citrate was quantitatively converted to large amounts of succinate and small amounts of α-ketoglutarate. (3) Addition of oxaloacetate in the absence of oxygen led to production of a large amount of citrate; the amount was increased if glucose was also added.
Other workers had found the following pathway in similar muscle tissue preparations:
- Based only on the data presented in this problem, what is the order of the intermediates in the citric acid cycle? How does this compare with Figure 16-7? Explain your reasoning.
- Why was it important to show the quantitative conversion of citrate to α-ketoglutarate?
The Krebs and Johnson article also contains other data that filled in most of the missing components of the cycle. The only component left unresolved was the molecule that reacted with oxaloacetate to form citrate.
Reference
- Krebs, H.A., and W.A. Johnson. 1937. The role of citric acid in intermediate metabolism in animal tissues. Enzymologia 4:148–156. Reprinted in FEBS Lett. 117(Suppl.):K2–K10, 1980.