Chapter Review
KEY TERMS
Terms in bold are defined in the glossary.
- micelle
- bilayer
- vesicle
- fluid mosaic model
- lipid transfer protein
- integral proteins
- peripheral proteins
- amphitropic proteins
- monotopic
- bitopic
- polytopic
- hydropathy index
- β barrel
- porin
- positive-inside rule
- GPI-anchored protein
- flippases
- floppases
- scramblases
- FRAP
- microdomains
- rafts
- caveolae
- caveolin
- BAR domain
- septins
- fusion protein
- v-SNAREs
- t-SNAREs
- integrin
- selectin
- simple diffusion
- membrane potential
- electrochemical gradient
- electrochemical potential
- transporters
- passive transport
- active transport
- ion channels
- amphipathic
- electroneutral
- cotransport
- antiport
- symport
- uniport
- electrogenic
- P-type ATPases
- SERCA pump
- ATPase
- V-type ATPases
- F-type ATPases
- ATP synthase
- ABC transporters
- multidrug transporters
- -glucose symporter
- ionophore
- aquaporins (AQPs)
- ligand-gated channel
- voltage-gated ion channel
- patch-clamping
Problems
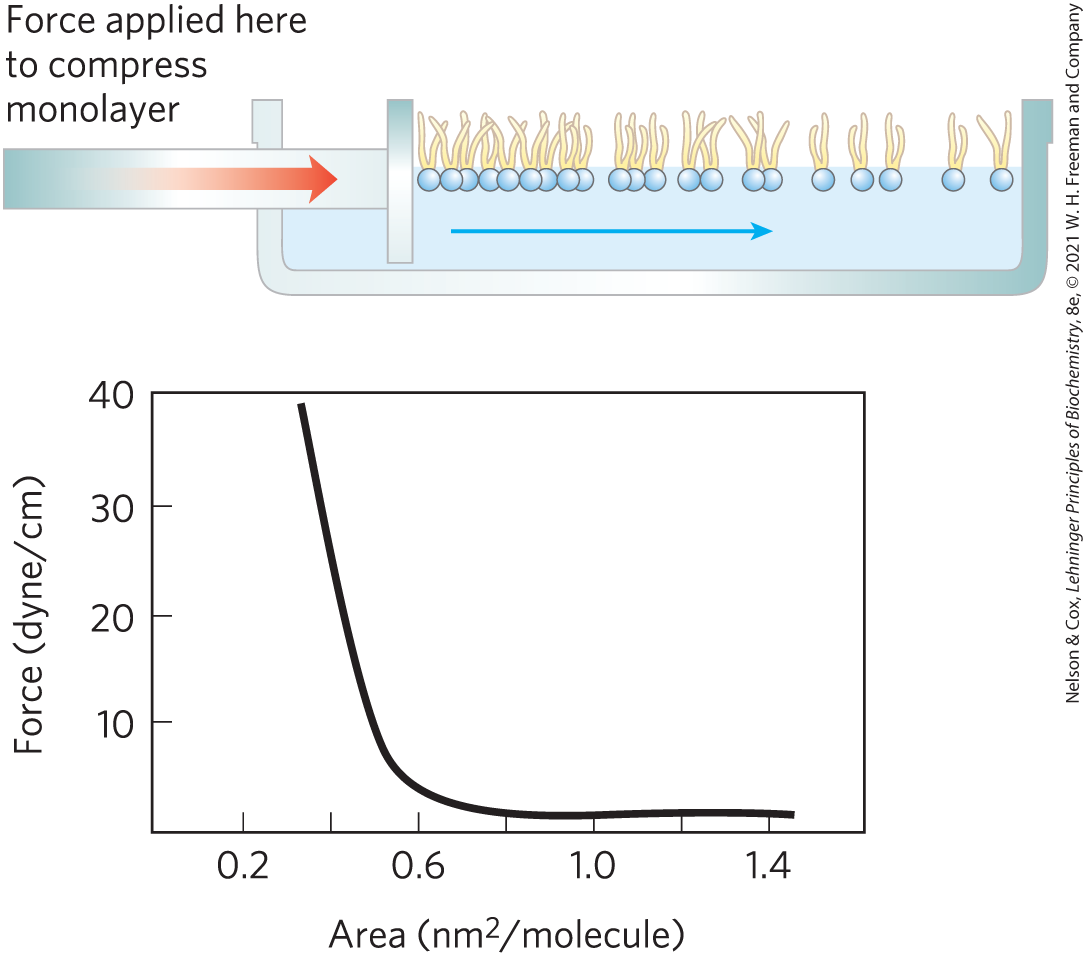
1. Determining the Cross-Sectional Area of a Lipid Molecule When phospholipids are layered gently onto the surface of water, they orient at the air-water interface with their head groups in the water and their hydrophobic tails in the air. The experimental apparatus shown can be used to progressively reduce the surface area available to a layer of lipids. By measuring the force necessary to push the lipids together, we can determine when the molecules are packed tightly in a continuous monolayer; as that area is approached, the force needed to further reduce the surface area increases sharply, as shown in the graph. How would you use this apparatus to determine the average area occupied by a single lipid molecule in the monolayer?
2. Properties of Lipids and Lipid Bilayers Lipid bilayers form when phospholipids are suspended in water. The edges of these sheets close upon each other and undergo self-sealing to form vesicles (liposomes).
- What properties of lipids are responsible for this property of bilayers? Explain.
- What are the consequences of this property for the structure of biological membranes?
3. Length of a Fatty Acid Molecule The carbon–carbon bond distance for single-bonded carbons, such as those in a saturated fatty acyl chain, is about 1.5 Å. Estimate to one significant figure the length of a single molecule of palmitate in its fully extended form. If two molecules of palmitate were placed end to end, how would their total length compare with the thickness of the lipid bilayer in a biological membrane?
4. Membrane Proteins What are the three main categories of membrane proteins, and how are they distinguished experimentally?
5. Location of a Membrane Protein Treatment of disrupted erythrocyte membranes with a concentrated salt solution released an unknown membrane protein, X. Proteolytic enzymes cleaved X into fragments. In additional experiments, intact erythrocytes were treated with proteolytic enzymes, washed, then disrupted. Extraction of membrane components yielded intact X. What do these observations indicate about the location of X in the plasma membrane? Do the properties of X resemble those of an integral membrane protein or a peripheral membrane protein?
6. Predicting Membrane Protein Topology from Sequence You have cloned the gene for a human erythrocyte protein, which you suspect is a membrane protein. You deduce the amino acid sequence of the protein from the nucleotide sequence of the gene. From this sequence alone, how would you evaluate the possibility that the protein is an integral protein? Suppose the protein proves to be an integral protein with one transmembrane segment. Suggest biochemical or chemical experiments that might allow you to determine whether the protein is oriented with the amino terminus on the outside of the cell or on the inside of the cell.
7. Surface Density of a Membrane Protein E. coli can be induced to make about 10,000 copies of the lactose transporter ( 31,000) per cell. Assume that E. coli is a cylinder 1 µm in diameter and 2 µm long. The diameter of the lactose transporter is approximately 6 nm. What fraction of the plasma membrane surface is occupied by the lactose transporter molecules? Explain how you arrived at this conclusion.
8. Molecular Species in the Plasma Membrane The plasma membrane of E. coli is about 75% protein and 25% phospholipid by weight. How many molecules of membrane lipid are present for each molecule of membrane protein? Assume an average protein of 50,000 and an average phospholipid of 750. What more would you need to know to estimate the fraction of the membrane surface that is covered by lipids?
9. Temperature Dependence of Lateral Diffusion The experiment described in Figure 11-19 was performed at 37 °C. If the experiment were carried out at 10 °C, what effect would you expect on the rate of diffusion? Why?
10. Membrane Self-Sealing Cellular membranes are self-sealing — if they are punctured or disrupted mechanically, they quickly and automatically reseal. What properties of membranes are responsible for this important feature?
11. Lipid Melting Temperatures Membrane lipids in tissue samples obtained from different parts of a reindeer’s leg have different fatty acid compositions. Membrane lipids from tissue near the hooves contain a larger proportion of unsaturated fatty acids than those from tissue in the upper leg. What is the significance of this observation?
12. Flip-Flop Diffusion What is the physical explanation for the very slow movement of membrane phospholipids from one leaflet of a biological membrane to the other? What factors influence this rate?
13. Bilayer Asymmetry The inner leaflet (monolayer) of the human erythrocyte membrane consists predominantly of phosphatidylethanolamine and phosphatidylserine. The outer leaflet consists predominantly of phosphatidylcholine and sphingomyelin. Although the phospholipid components of the membrane can diffuse in the fluid bilayer, this sidedness is preserved at all times. How?
14. Scramblase and Flippase Explain the difference between the scramblase enzymes and flippase enzymes based on the membranes with which they are associated, the symmetry of these membranes, and their energy requirements.
15. Membrane Permeability At pH 7, tryptophan crosses a lipid bilayer at about one-thousandth the rate of indole, a closely related compound:
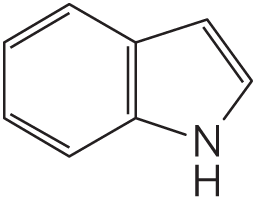
Suggest an explanation for this observation.
16. Glucose Transporters A cell biologist working with cultured cells from intestinal epithelium finds that the cells take up glucose from the growth medium 10 times faster when the glucose concentration is 5 mm than when it is 0.2 mm. She also finds that glucose uptake requires in the growth medium. What can you say about the glucose transporter in these cells?
17. Use of the Helical Wheel Diagram A helical wheel is a two-dimensional representation of a helix, a view along its central axis. Use the helical wheel diagram shown here to determine the distribution of amino acid residues in a helical segment with the sequence –Val–Asp–Arg–Val–Phe–Ser–Asn–Val–Cys–Thr–His–Leu–Lys–Thr–Leu–Gln–Asp–Lys–
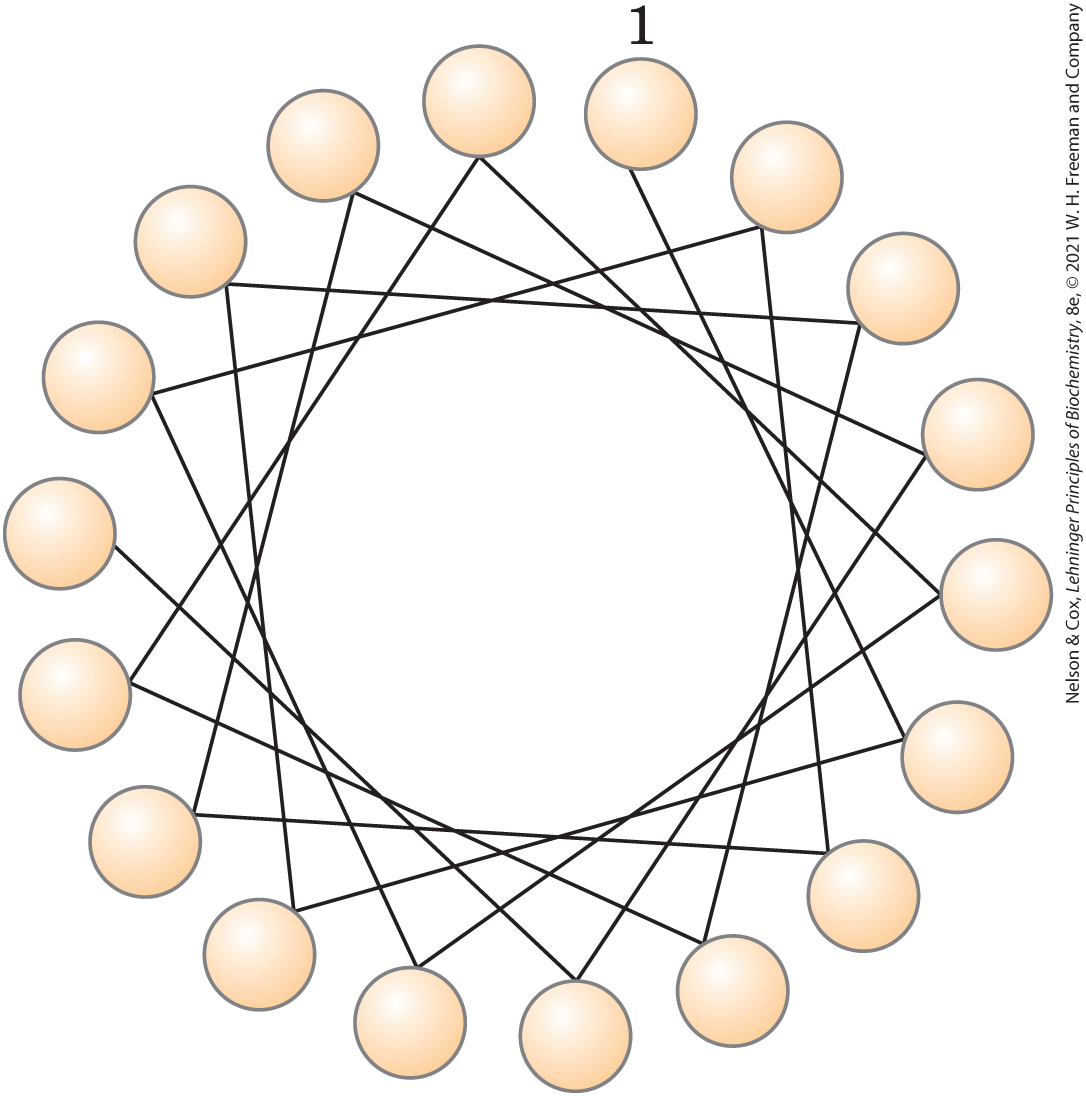
What can you say about the surface properties of this helix? How would you expect the helix to be oriented in the tertiary structure of an integral membrane protein?
18. Synthesis of Gastric Juice: Energetics Parietal cells acidify gastric juice (pH 1.5) by pumping HCl from blood plasma (pH 7.4) into the stomach. How much ATP (in moles) is required to pump a mole of protons () against this concentration gradient? The free-energy change for ATP hydrolysis under cellular conditions is about –58 kJ/mol. Ignore the effects of the transmembrane electrical potential.
19. Electrogenic Transporters A single-cell organism, Paramecium, is large enough to allow the insertion of a microelectrode, permitting the measurement of the electrical potential between the inside of the cell and the surrounding medium (the membrane potential). The measured membrane potential is −50 mV (inside negative) in a living cell. What would happen if you added valinomycin to the surrounding medium, which contains and ?
20. Energetics of the ATPase For a typical vertebrate cell with a membrane potential of −0.070 V (inside negative), what is the free-energy change for transporting 1 mol of from the cell into the blood at 37 °C? Assume the concentration is 12 mm inside the cell and 145 mm in blood plasma.
21. Action of Ouabain on Kidney Tissue Ouabain specifically inhibits the ATPase activity of animal tissues but is not known to inhibit any other enzyme. When ouabain is added to thin slices of living kidney tissue, it inhibits oxygen consumption by 66%. Why? What does this observation tell us about the use of respiratory energy by kidney tissue?
22. Digoxin to Inhibit ATPase The exchanger expressed in cardiac myocytes is a bidirectional antiporter protein that removes calcium from the cytoplasm by exchanging it with sodium. Cardiac myocytes also express the ATPase. Suppose that a ATPase inhibitor (digoxin) is added to cardiac myocytes. Using your knowledge of the relative concentrations of ions (intracellular versus extracellular) and the important role of the ATPase in maintaining the electrochemical gradient, what change would you expect in the intracellular []? Why?
23. Energetics of Symport Suppose you determined experimentally that a cellular transport system for glucose, driven by symport of , could accumulate glucose to concentrations 25 times greater than in the external medium, while the external [] was only 10 times greater than the intracellular []. Would this violate the laws of thermodynamics? If not, how could you explain this observation?
24. Labeling the Lactose Transporter A bacterial lactose transporter, which is highly specific for lactose, contains a Cys residue that is essential to its transport activity. Covalent reaction of N-ethylmaleimide (NEM) with this Cys residue irreversibly inactivates the transporter. A high concentration of lactose in the medium prevents inactivation by NEM, presumably by sterically protecting the Cys residue, which is in or near the lactose-binding site. You know nothing else about the transporter protein. Suggest an experiment that might allow you to determine the of this Cys-containing transporter polypeptide.
25. Transport Types You have just discovered a new l-alanine transporter in liver cells (hepatocytes). Poisoning hepatocytes with cyanide (which blocks ATP synthesis) reduces alanine transport by 90%. Tenfold reduction in extracellular [] has no immediate effect on alanine transport. How would you use these observations to decide whether the alanine transporter is passive or active, primary or secondary?
26. Intestinal Uptake of Leucine You are studying the uptake of l-leucine by epithelial cells of the mouse intestine. Measurements of the rates of uptake of l-leucine and several of its analogs, with and without in the assay buffer, yield the results given in the table. What can you conclude about the properties and mechanism of the leucine transporter? Would you expect l-leucine uptake to be inhibited by ouabain (see problem 21)?
Substrate Uptake in presence of Uptake in absence of l-Leucine 420 0.24 23 0.2 d-Leucine 310 4.7 5 4.7 l-Valine 225 0.31 19 0.31 27. Ion Channel Selectivity Potassium channels consist of four subunits that form a channel just wide enough for ions to pass through. Although ions are smaller ( 23, radius 0.95 Å) than ions ( 39, radius 1.33 Å), the potassium channels in the bacterium Streptomyces lividans transport 104 times more ions than ions. What prevents ions from passing through potassium channels?
28. Effect of an Ionophore on Active Transport Consider the leucine transporter described in Problem 26. Would and/or change if you added a ionophore to the assay solution containing ? Explain.
Biochemistry Online
29. Predicting Membrane Protein Topology I Online bioinformatics tools make hydropathy analysis easy if you know the amino acid sequence of a protein. At the Protein Data Bank (www.rcsb.org), the Protein Feature View displays additional information about a protein gleaned from other databases, such as UniProt and SCOP2. A simple graphical view of a hydropathy plot created using a window of 15 residues shows hydrophobic regions in red and hydrophilic regions in blue.
- Looking only at the displayed hydropathy plots in the Protein Feature View, what predictions would you make about the membrane topology of these proteins: glycophorin A (PDB ID 1AFO), myoglobin (PDB ID 1MBO), and aquaporin (PDB ID 2B6O)?
- Now, refine your information using the ProtScale tools at the ExPASy bioinformatics resource portal. Each of the PDB Protein Feature Views was created with a UniProt Knowledgebase ID. For glycophorin A, the UniProtKB ID is P02724; for myoglobin, P02185; and for aquaporin, Q6J8I9. Go to the ExPASy portal (http://web.expasy.org/protscale) and select the Kyte & Doolittle hydropathy analysis option, with a window of 7 amino acids. Enter the UniProtKB ID for aquaporin (Q6J8I9, which you can also get from the PDB’s Protein Feature View page), then select the option to analyze the complete chain (residues 1 to 263). Use the default values for the other options and click Submit to get a hydropathy plot. Save a GIF image of this plot. Now repeat the analysis using a window of 15 amino acids. Compare the results for the 7-residue and 15-residue window analyses. Which window size gives you a better signal-to-noise ratio?
- Under what circumstances would it be important to use a narrower window?
30. Predicting Membrane Protein Topology II The epinephrine receptor in animal cells is an integral membrane protein ( 64,000) that is believed to have seven transmembrane α-helical regions.
- Show that a protein of this size is capable of spanning the membrane seven times.
- Given the amino acid sequence of this protein, how would you predict which regions of the protein form the membrane-spanning helices?
- Go to the Protein Data Bank (www.rcsb.org). Use the PDB identifier 1DEP to retrieve the data page for a portion of the β-adrenergic receptor (one type of epinephrine receptor) isolated from turkey. Using JSmol to explore the structure, predict whether this portion of the receptor is located within the membrane or at the membrane surface. Explain your answer. Now use the Protein Feature View to see the hydrophobicity analysis of the sequence. Does this support your answer?
- Retrieve the data for a portion of another receptor, the acetylcholine receptor of neurons and myocytes, using the PDB identifier 1A11. As in (c), predict where this portion of the receptor is located and explain your answer.
Data Analysis Problem
31. The Fluid Mosaic Model of Biological Membrane Structure Figure 11-3 shows the currently accepted fluid mosaic model of biological membrane structure. This model was presented in detail in a review article by S. J. Singer in 1971. In the article, Singer presented the three models of membrane structure that had been proposed up to that time:
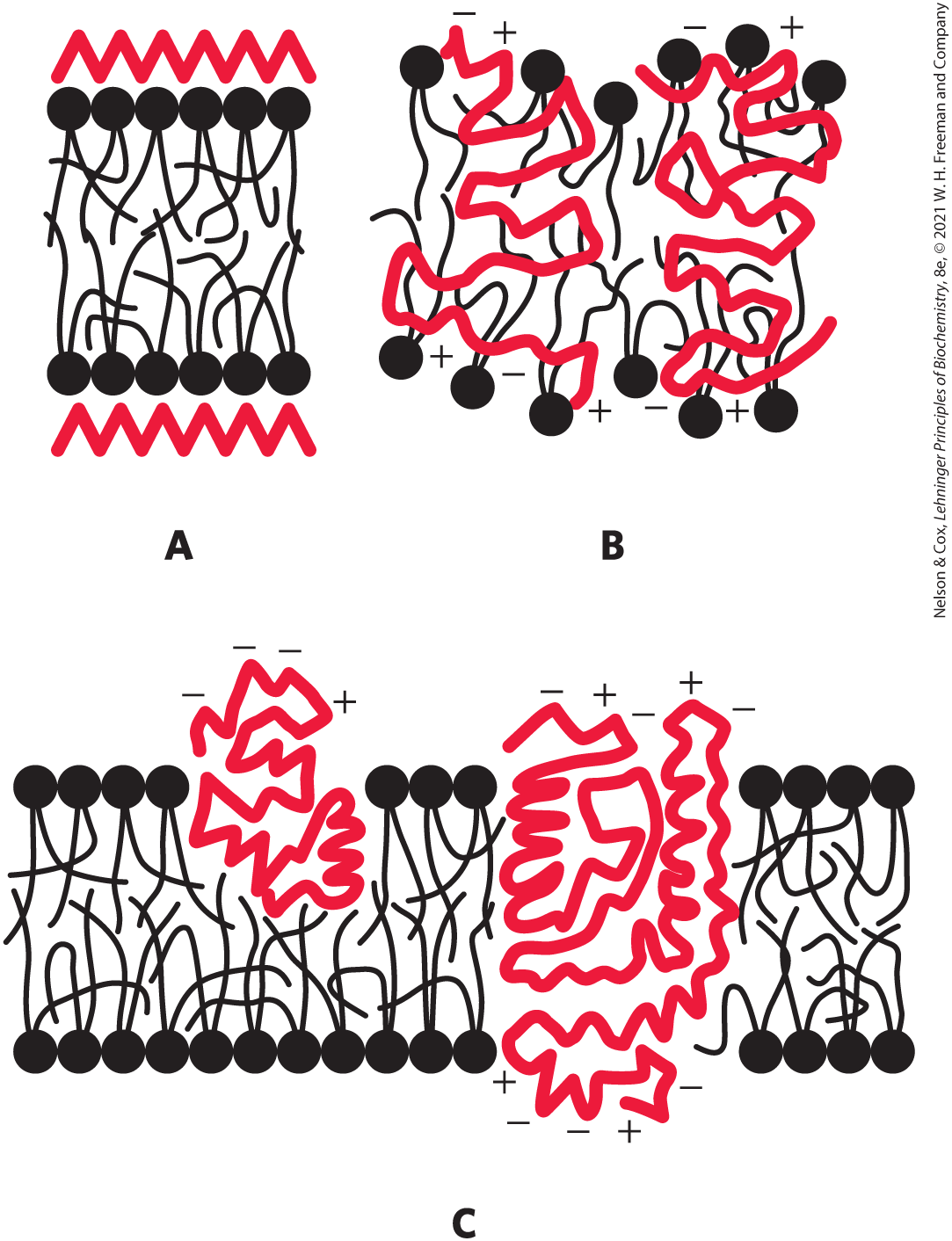
A. The Davson-Danielli-Robertson Model. This was the most widely accepted model in 1971, when Singer’s review was published. In this model, the phospholipids are arranged as a bilayer. Proteins are found on both surfaces of the bilayer, attached to it by ionic interactions between the charged head groups of the phospholipids and charged groups of the proteins. Crucially, there is no protein in the interior of the bilayer.
B. The Benson Lipoprotein Subunit Model. Here the proteins are globular and the membrane is a protein-lipid mixture. The hydrophobic tails of the lipids are embedded in the hydrophobic parts of the proteins. The lipid head groups are exposed to the solvent. There is no lipid bilayer.
C. The Lipid–Globular Protein Mosaic Model. This is the model shown in Figure 11-3. The lipids form a bilayer and proteins are embedded in it, some extending through the bilayer and others not. Proteins are anchored in the bilayer by interactions between the hydrophobic tails of the lipids and hydrophobic portions of the protein.
For the data given below, consider how each piece of information aligns with each of the three models of membrane structure. Which model(s) are supported, which are not supported, and what reservations do you have about the data or their interpretation? Explain your reasoning.
- When cells were fixed, stained with osmium tetroxide, and examined in the electron microscope, the membranes showed a “railroad track” appearance, with two dark-staining lines separated by a light space.
- The thickness of membranes in cells fixed and stained in the same way was found to be 5 to 9 nm. The thickness of a “naked” phospholipid bilayer, without proteins, was 4 to 4.5 nm. The thickness of a single monolayer of proteins was about 1 nm.
- Singer wrote in his article: “The average amino acid composition of membrane proteins is not distinguishable from that of soluble proteins. In particular, a substantial fraction of the residues is hydrophobic” (p. 165).
- As described in Problems 1 and 2 of this chapter, researchers had extracted membranes from cells, extracted the lipids, and compared the area of the lipid monolayer with the area of the original cell membrane. The interpretation of the results was complicated by the issue illustrated in the graph of Problem 1: the area of the monolayer depended on how hard it was pushed. With very light pressures, the ratio of monolayer area to cell membrane area was about 2.0. At higher pressures — thought to be more like those found in cells — the ratio was substantially lower.
- Circular dichroism spectroscopy uses changes in polarization of UV light to make inferences about protein secondary structure (see Fig. 4-9). On average, this technique showed that membrane proteins have a large amount of α helix and little or no β sheet. This finding was consistent with most membrane proteins having a globular structure.
- Phospholipase C is an enzyme that removes the polar head group (including the phosphate) from phospholipids. In several studies, treatment of intact membranes with phospholipase C removed about 70% of the head groups without disrupting the “railroad track” structure of the membrane.
- Singer described in his article a study in which “a glycoprotein of molecular weight about 31,000 in human red blood cell membranes is cleaved by tryptic treatment of the membranes into soluble glycopeptides of about 10,000 molecular weight, while the remaining portions are quite hydrophobic” (p. 199). Trypsin treatment did not cause gross changes in the membranes, which remained intact.
Singer’s review also included many more studies in this area. In the end, though, the data available in 1971 did not conclusively prove Model C was correct. As more data have accumulated, this model of membrane structure has been accepted by the scientific community.
Reference
- Singer, S.J. 1971. The molecular organization of biological membranes. In Structure and Function of Biological Membranes (L.I. Rothfield, ed.), pp. 145–222. New York: Academic Press.