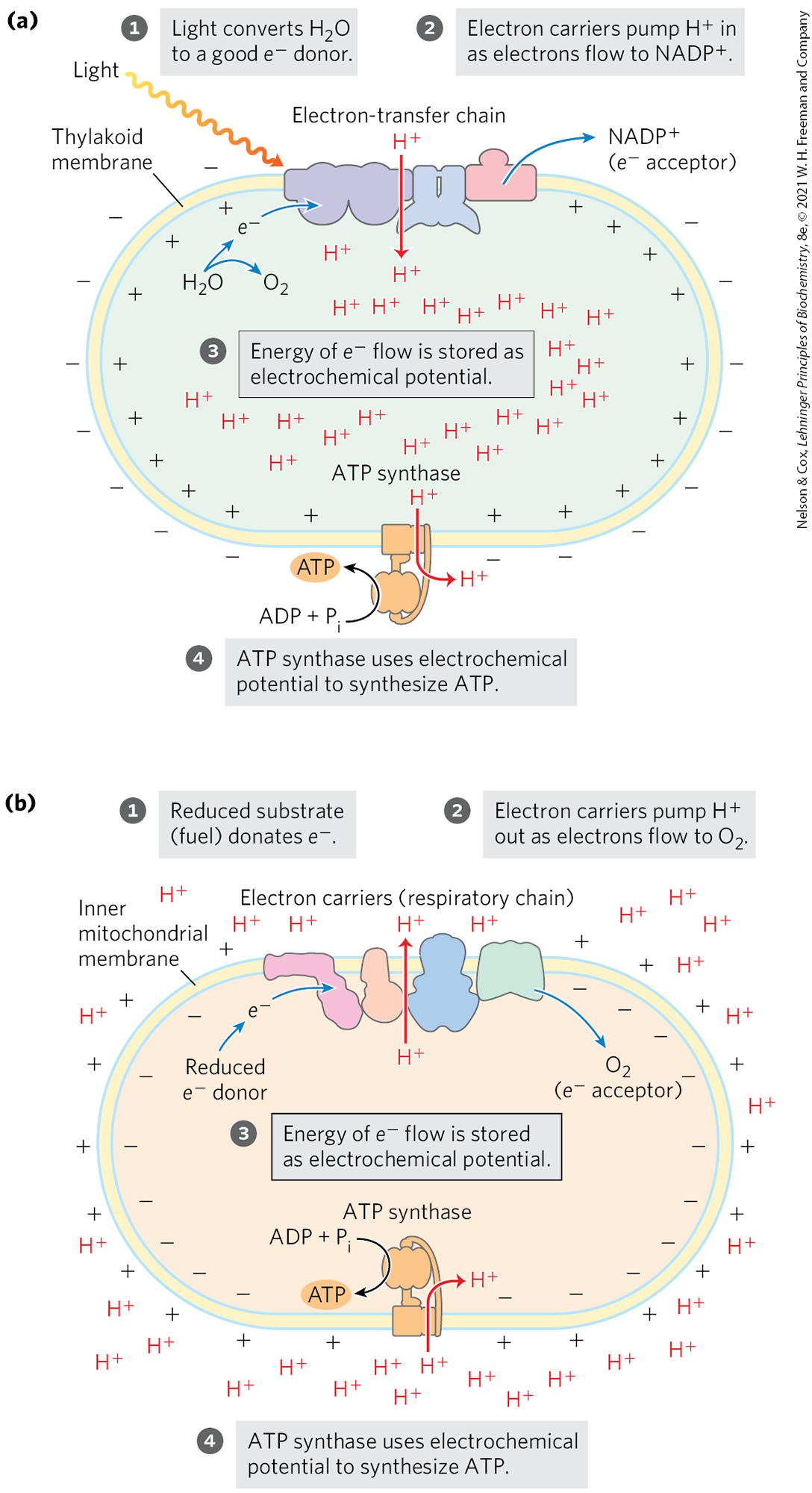
Photophosphorylation (ATP synthesis driven by light) resembles oxidative phosphorylation in that electron flow through a series of membrane carriers is coupled to proton pumping, producing the proton motive force that powers ATP formation. The processes are compared in Figure 20-2. In oxidative phosphorylation, the electron donor is NADH and the ultimate electron acceptor is , forming . In photophosphorylation, electrons flow in the opposite direction: is the electron donor and NADPH is formed. How is this endergonic process possible?
FIGURE 20-2 The chemiosmotic mechanism for ATP synthesis in chloroplasts and mitochondria. (a) Movement of electrons through a chain of membrane-bound carriers in the chloroplast membrane is driven by the energy of photons absorbed by the green pigment chlorophyll. Electron flow leads to the movement of protons and positive charge across the membrane, creating an electrochemical potential. This electrochemical potential drives ATP synthesis by the membrane-bound ATP synthase, which is fundamentally similar in structure and mechanism to (b) the mitochondrial machinery for oxidative phosphorylation of mitochondria. In mitochondria, the force that moves electrons through the complexes is a large difference in the reduction potentials of electron donor and acceptor. In both systems, the energy made available by electron transfer is captured as a transmembrane proton gradient, which drives ATP synthesis by an ATP synthase.
Water is a poor donor of electrons; its standard reduction potential is 0.816 V, compared with for NADH, a good electron donor. Photosynthesis requires the input of energy in the form of light to create a good electron donor and a good electron acceptor. Electrons flow from the electron donor through a series of membrane-bound carriers, including cytochromes, quinones, and iron-sulfur proteins, while protons are pumped across a membrane to create an electrochemical potential. Electron transfer and proton pumping are catalyzed by a membrane complex that is homologous in structure and function to Complex III of mitochondria. The electrochemical potential so produced is the driving force for ATP synthesis from ADP and , catalyzed by a membrane-bound ATP synthase complex closely similar to that of mitochondria and bacteria.
Chloroplasts Are the Site of Light-Driven Electron Flow and Photosynthesis in Plants
In photosynthetic eukaryotic cells, both the light-dependent and the -assimilation reactions take place in chloroplasts (Fig. 20-3), organelles that are variable in shape and generally a few micrometers in diameter. Like mitochondria, chloroplasts are surrounded by two membranes: an outer membrane that is permeable to small molecules and ions, and an impermeable inner membrane that bears specific transporters for a variety of ions and metabolites. The space enclosed by the inner membrane is called the stroma in chloroplasts and is analogous to the mitochondrial matrix; it is an aqueous phase containing most of the soluble enzymes required for the -assimilation reactions. Throughout the stroma is a highly organized set of topologically continuous internal membranes, forming a single compartment or lumen. This complex membrane system forms flattened sacks called thylakoids. Granal thylakoids are disk-like pouches arranged in stacks; they are connected by stromal thylakoids, which are flatter and spiral around a stack of grana. The thylakoid membranes provide a large area for the machinery of photophosphorylation — the photosynthetic pigments and enzyme complexes that carry out the light-dependent reactions and ATP synthesis. Traffic across these membranes is also mediated by specific transporters.
FIGURE 20-3 Chloroplast structure. (a) Schematic diagram. (b) Colorized electron micrograph at high magnification, showing the highly organized thylakoid membrane system.
In 1937, Robert Hill found that when leaf extracts containing chloroplasts were illuminated, they (1) evolved and (2) reduced a nonbiological electron acceptor added to the medium, according to the Hill reaction
where A is an artificial electron acceptor, or Hill reagent. One Hill reagent, the dye 2,6-dichlorophenolindophenol, is blue when oxidized (A) and colorless when reduced , making the reaction easy to follow.
When a leaf extract supplemented with the dye was illuminated, the blue dye became colorless and was evolved. In the dark, no evolution or dye reduction took place. This was the first evidence that absorbed light energy causes electrons to flow from some electron donor (now known to be ) to an electron acceptor. Moreover, Hill found that was neither required nor reduced to a stable form under these conditions; production could be dissociated from reduction. Several years later, Severo Ochoa showed that is the biological electron acceptor in chloroplasts, according to the equation
To understand this photochemical process, we must first consider the more general topic of the effects of light absorption on molecular structure.
Visible light is electromagnetic radiation of wavelengths 400 to 700 nm, a small part of the electromagnetic spectrum (Fig. 20-4), ranging from violet to red. The energy of a single photon (a quantum of light) is greater at the violet end of the spectrum than at the red end; shorter wavelength (and higher frequency) corresponds to higher energy. The energy, E, in a single photon of visible light is given by the Planck equation:
where h is Planck’s constant , ν is the frequency of the light in cycles/s, c is the speed of light , and λ is the wavelength of the light in meters. The energy of a photon of visible light ranges from 150 kJ/einstein for red light to ∼300 kJ/einstein for violet light.
FIGURE 20-4 Electromagnetic radiation. The spectrum of electromagnetic radiation, and the energy of photons in the visible range. One einstein is photons.
WORKED EXAMPLE 20-1 Energy of a Photon
The light used by vascular plants for photosynthesis has a wavelength of about 700 nm. Calculate the energy in a “mole” of photons (an einstein) of light of this wavelength, and compare this with the energy needed to synthesize a mole of ATP.
SOLUTION:
The energy in a single photon is given by the Planck equation. At a wavelength of , the energy of a photon is
An einstein of light is Avogadro’s number of photons ; thus the energy of one einstein of photons at 700 nm is given by
So, a “mole” of photons of red light has about five times the energy needed to produce a mole of ATP from ADP and (30.5 kJ/mol).
When a photon is absorbed, an electron in the absorbing molecule (chromophore) is lifted to a higher energy level. This is an all-or-nothing event: to be absorbed, the photon must contain a quantity of energy, called a quantum, that exactly matches the energy of the electronic transition. A molecule that has absorbed a photon is in an excited state, which is generally unstable. An electron lifted into a higher-energy orbital usually returns rapidly to its lower-energy orbital; that is, the excited molecule decays to the stable ground state, giving up the absorbed quantum as light or heat or using it to do chemical work. Light emission accompanying decay of excited molecules, fluorescence, is always at a longer wavelength (lower energy) than that of the absorbed light (see Box 12-1). An alternative mode of decay, central to photosynthesis, involves direct transfer of excitation energy from an excited molecule to a neighboring molecule. Just as the photon is a quantum of light energy, so the exciton is a quantum of energy passed from an excited molecule to another molecule in a process called exciton transfer.
Chlorophylls Absorb Light Energy for Photosynthesis
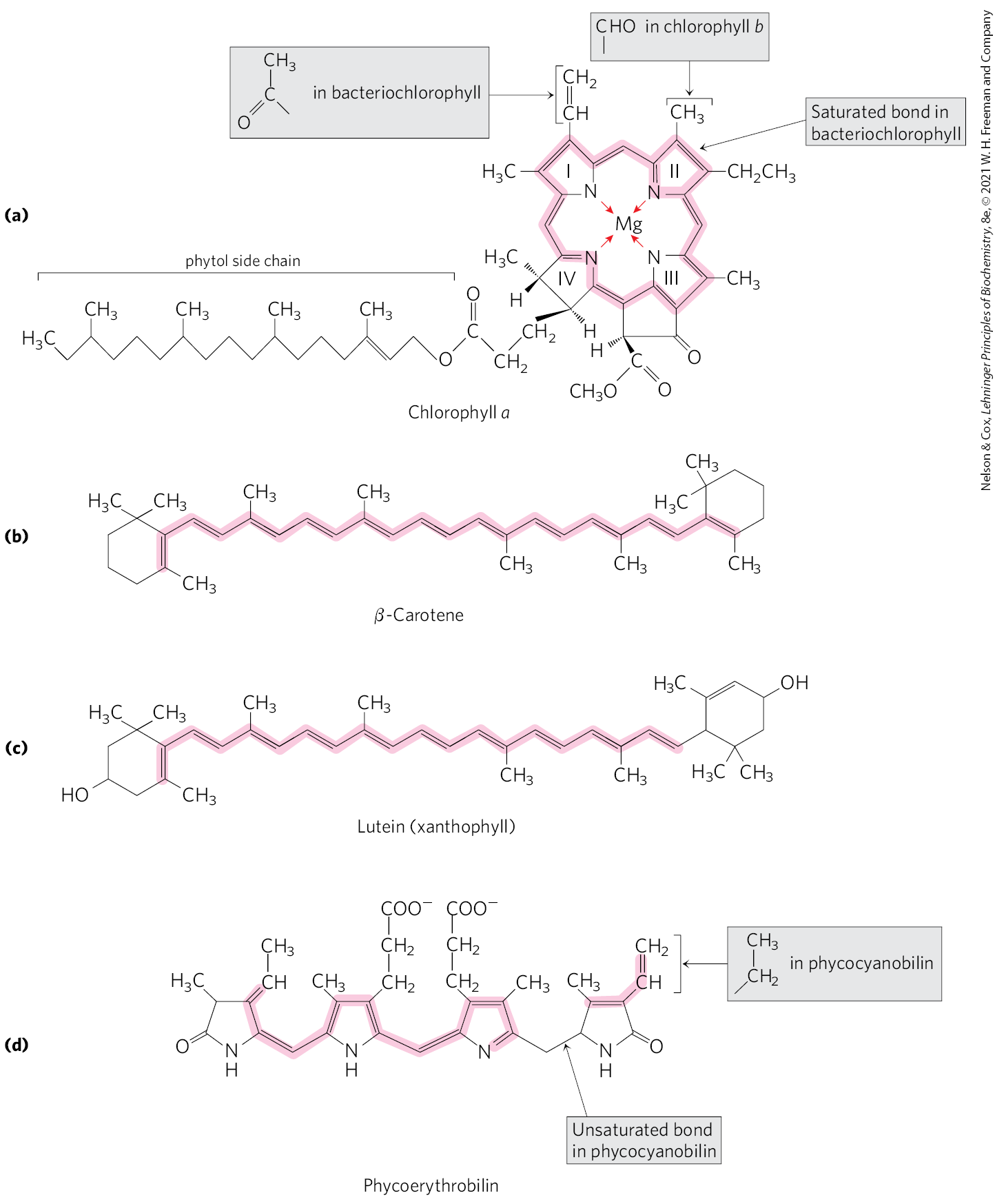
The most important light-absorbing pigments in the thylakoid membranes are the chlorophylls, green pigments with polycyclic, planar structures resembling the protoporphyrin of hemoglobin, except that , not , occupies the central position (Fig. 20-5a; compare to Fig. 5-1). The four inward-oriented nitrogen atoms of chlorophyll are coordinated with the . All chlorophylls have a long phytol side chain, esterified to a carboxyl-group substituent in ring IV, and chlorophylls also have a fifth five-membered ring not present in heme.
FIGURE 20-5 Primary and secondary photopigments. (a) Chlorophylls a and b and bacteriochlorophyll are the primary gatherers of light energy. (b) β-Carotene (a carotenoid) and (c) lutein (a xanthophyll) are accessory pigments in plants. (d) Phycoerythrobilin and phycocyanobilin (phycobilins) are accessory pigments in cyanobacteria and red algae. The conjugated bond systems in these molecules (alternating single and double bonds, shaded) have delocalized electrons that are easily excited by photons with the wavelengths of visible light.
The heterocyclic five-ring system that surrounds the has an extended polyene structure, with alternating single and double bonds. Such polyenes characteristically show strong absorption in the visible region of the spectrum (Fig. 20-6); the chlorophylls have unusually high molar extinction coefficients (see Box 3-1) and are therefore particularly well-suited for absorbing visible light during photosynthesis.
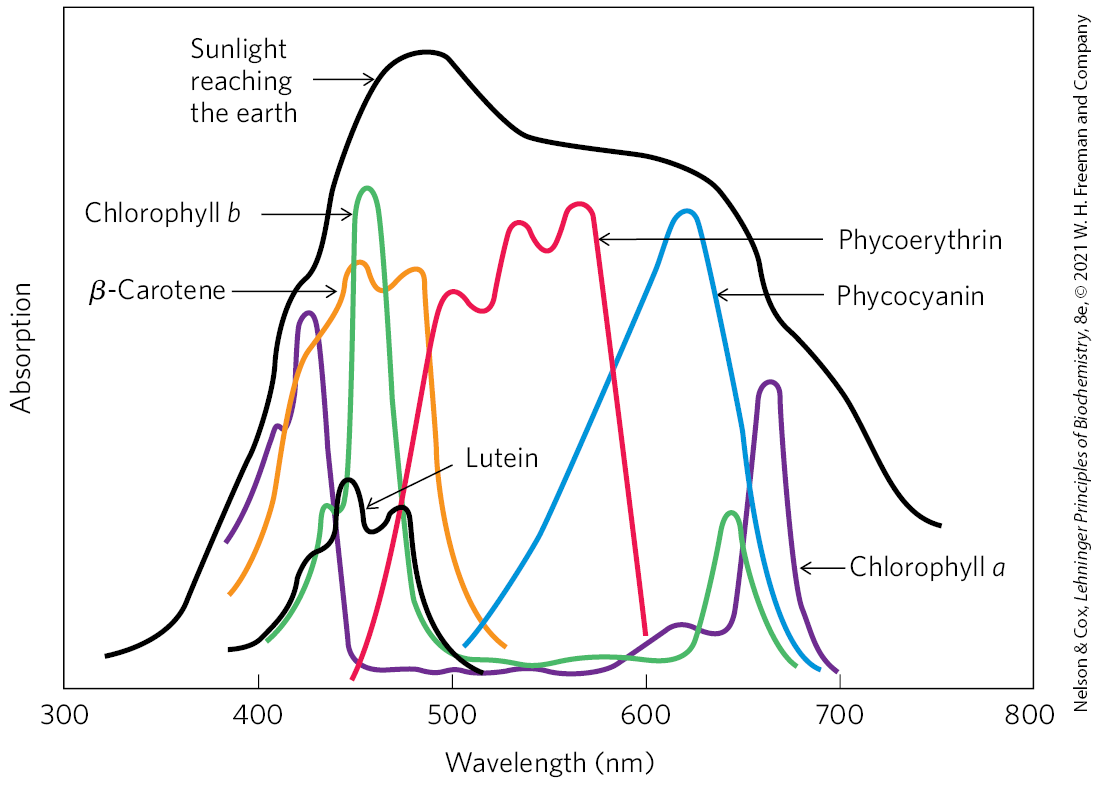
FIGURE 20-6 Absorption of visible light by photopigments. Plants are green because their pigments absorb light from the red and blue regions of the spectrum, leaving primarily green light to be reflected. Compare the absorption spectra of the pigments with the spectrum of sunlight reaching the earth’s surface; the combination of chlorophylls (a and b) and accessory pigments enables plants to harvest most of the energy available in sunlight. The relative amounts of chlorophylls and accessory pigments are characteristic of a particular plant species. Variation in the proportions of these pigments is responsible for the range of colors of photosynthetic organisms, from the deep blue-green of spruce needles, to the greener green of maple leaves, to the red, brown, or purple color of some species of multicellular algae and the leaves of some foliage plants favored by gardeners.
Chloroplasts always contain both chlorophyll a and chlorophyll b (Fig. 20-5a). Although both are green, their absorption spectra are sufficiently different (Fig. 20-6) that they complement each other’s range of light absorption in the visible region. Most plants contain about twice as much chlorophyll a as chlorophyll b. The chlorophyll in cyanobacteria differs only slightly from those of plants.
In addition to chlorophylls, thylakoid membranes of plants contain secondary light-absorbing pigments, or accessory pigments, called carotenoids. Carotenoids may be yellow, red, or purple. The two most prominent in plant leaves are β-carotene, a red-orange isoprenoid, and the yellow carotenoid lutein (Fig. 20-5b, c). Cyanobacteria and red algae use the accessory pigments phycocyanobilin and phycoerythrobilin (Fig. 20-5d). Accessory pigments absorb light at wavelengths not absorbed by the chlorophylls (Fig. 20-6) and thus are supplementary light receptors. They also protect downstream components from a highly reactive form of oxygen (singlet oxygen) that is formed when intense light exceeds the system’s capacity to accept electrons.
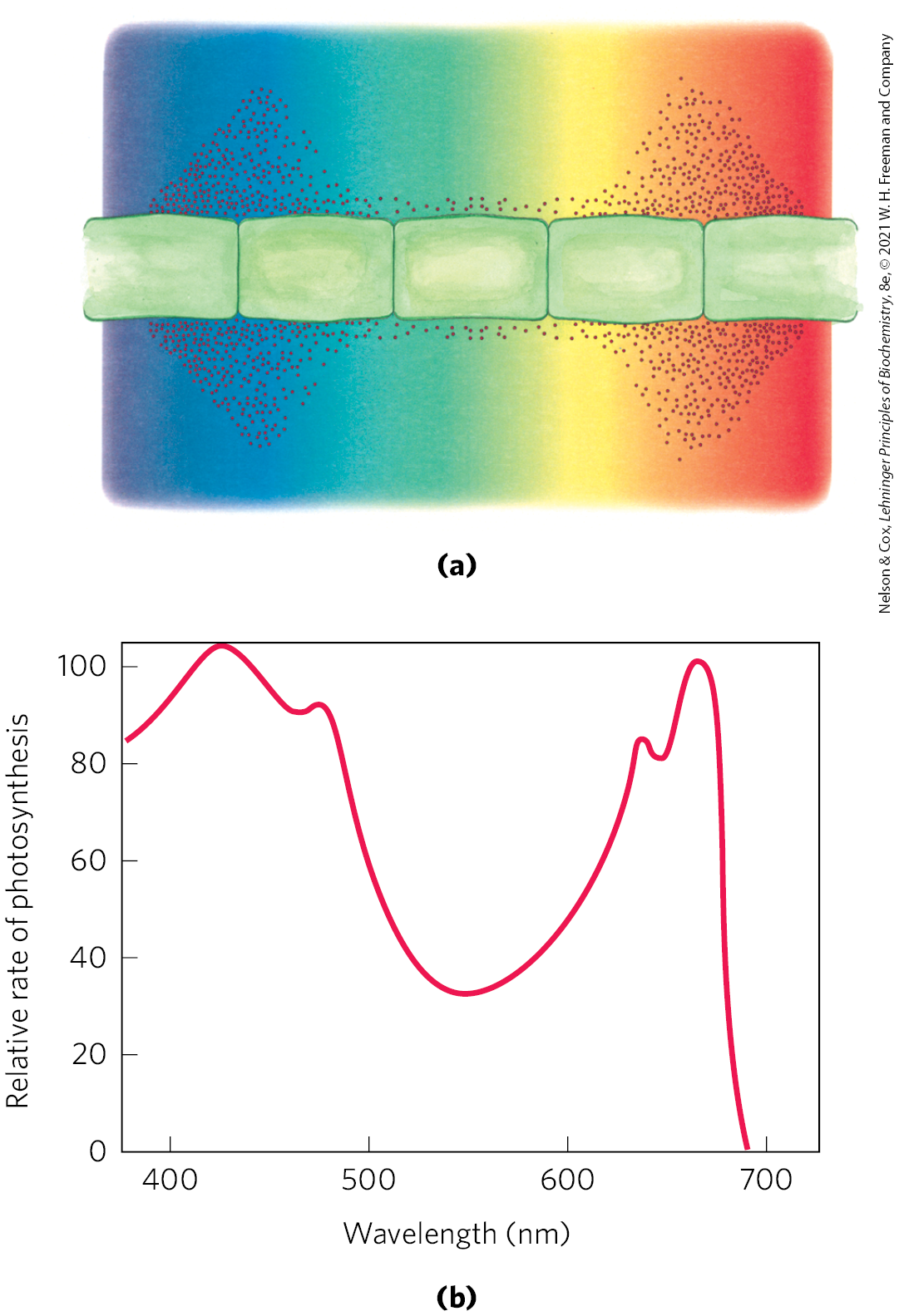
Experimental determination of the effectiveness of light of different colors in promoting photosynthesis yields an action spectrum (Fig. 20-7), often useful in identifying the pigment primarily responsible for a biological effect of light. By capturing light in a region of the spectrum not used by other organisms, a photosynthetic organism can claim a unique ecological niche.
FIGURE 20-7 Two ways to determine the action spectrum for photosynthesis. (a) Results of a classic experiment performed by T. W. Engelmann in 1882 to determine the wavelength of light that is most effective in supporting photosynthesis. Engelmann placed cells of a filamentous photosynthetic alga on a microscope slide and illuminated them with light from a prism, so that one part of the algal filament received mainly blue light, another part yellow, another red. To determine which cells carried out photosynthesis most actively, Engelmann also placed on the microscope slide bacteria known to migrate toward regions of high concentration. After a period of illumination, the distribution of bacteria showed highest levels (produced by photosynthesis) in the regions illuminated with violet and red light. (b) Results of a similar experiment that used modern techniques (an oxygen electrode) for the measurement of production. An action spectrum, as shown here, describes the relative rate of photosynthesis for illumination with a constant number of photons of different wavelengths. An action spectrum is useful because, by comparison with absorption spectra (such as those in Fig. 20-6), it suggests which pigments can channel energy into photosynthesis.
Chlorophylls Funnel Absorbed Energy to Reaction Centers by Exciton Transfer
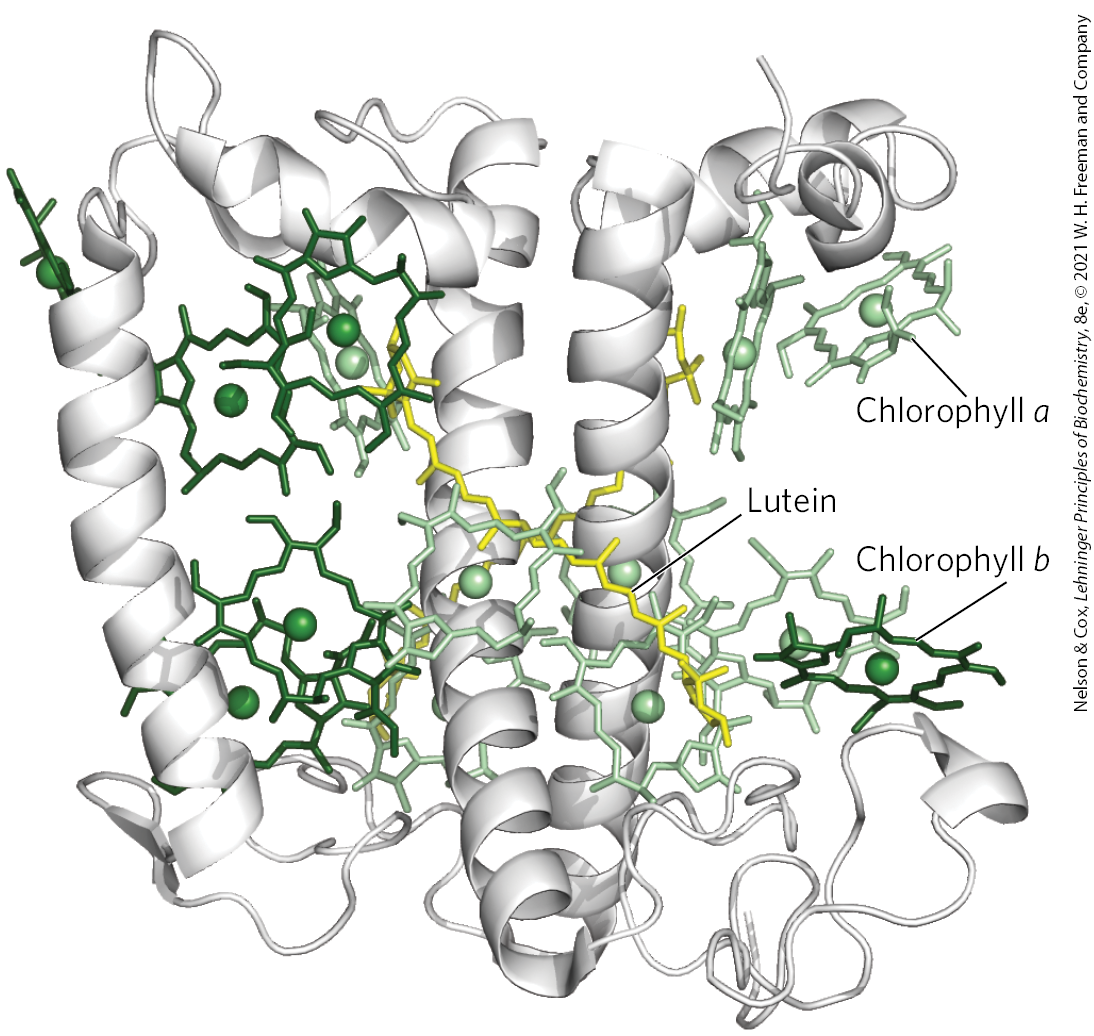
The light-absorbing pigments of thylakoid or bacterial membranes are arranged in functional arrays called photosystems. In spinach chloroplasts, for example, each photosystem contains about 200 chlorophyll and 50 carotenoid molecules. All the pigment molecules in a photosystem can absorb photons, but only one pair of chlorophyll molecules associated with the photochemical reaction center is specialized to transduce light into chemical energy. The other pigment molecules in a photosystem serve as antenna molecules. They absorb light energy and transmit it rapidly and efficiently to the reaction center (Fig. 20-8). Some chlorophylls are part of a core complex around the reaction center. Others form light-harvesting complexes (LHCs) around the periphery of the core complex. Chlorophyll and other pigments are always associated with specific binding proteins, which fix the chromophores in relation to each other, to other protein complexes, and to the membrane. For example, each monomer of the trimeric light-harvesting complex LHCII (Fig. 20-9) contains seven molecules of chlorophyll a, five of chlorophyll b, and two of lutein.
FIGURE 20-8 Organization of photosystems in the thylakoid membrane. Photosystems are tightly packed in the thylakoid membrane, with several hundred antenna chlorophylls and accessory pigments surrounding each reaction center. Absorption of a photon by any of the antenna chlorophylls leads to excitation of the reaction center by exciton transfer (red arrow).
FIGURE 20-9 The light-harvesting complex LHCII of the pea. The functional unit is a trimer, with 36 chlorophyll and 6 lutein molecules. Shown here is a monomer, viewed in the plane of the membrane, with its three transmembrane α-helical segments, seven chlorophyll a molecules (light green), five chlorophyll b molecules (dark green), and two molecules of lutein (yellow), which form an internal cross-brace. [Data from PDB ID 2BHW, J. Standfuss et al., EMBO J. 24:919, 2005.]
The chlorophyll molecules in light-harvesting complexes and other chlorophyll-binding proteins have light-absorption properties that are subtly different from those of free chlorophyll. When isolated chlorophyll molecules are excited by light, the absorbed energy is quickly released as fluorescence and heat; but when chlorophyll in intact leaves is excited by visible light (Fig. 20-10, step ), very little fluorescence is observed. Instead, the excited antenna chlorophyll transfers energy directly to a neighboring chlorophyll molecule, which becomes excited as the first molecule returns to its ground state (step ). This transfer of energy, exciton transfer, extends to a third, fourth, or subsequent neighbor, until one of a “special pair” of chlorophyll a molecules at the photochemical reaction center is excited (step ). The special pair of chlorophyll molecules, often designated , are held close enough to each other to share bonding orbitals, and to react as a single compound when excited. In this excited chlorophyll pair, an electron is promoted to a higher-energy orbital. This electron then passes to a nearby electron acceptor that is part of the photosynthetic electron-transfer chain, leaving the reaction-center chlorophyll pair with a missing electron (an “electron hole,” denoted by in Fig. 20-10) (step ). The electron acceptor acquires a negative charge in this transaction. The electron lost by the reaction-center chlorophyll pair is replaced by an electron from a neighboring electron-donor molecule (step ), which thereby becomes positively charged. In this way, excitation by light causes electric charge separation and initiates an oxidation-reduction chain.
FIGURE 20-10 Exciton and electron transfer. This generalized scheme shows conversion of the energy of an absorbed photon into separation of charges at the reaction center. Note that step may repeat between successive antenna molecules until the exciton reaches the special pair of chlorophylls in the reaction center. An asterisk (*) denotes the excited state of a molecule.
SUMMARY 20.1 Light Absorption
Photosynthesis takes place in plant chloroplasts, structures enclosed in double membranes and filled with an elaborate system of thylakoid membranes containing the photosynthetic machinery.
Chlorophyll molecules and other light-absorbing pigments are associated with proteins in light-harvesting complexes arrayed around photochemical reaction centers. The proteins are embedded in thylakoid membranes.
The many chlorophyll molecules that surround the reaction center serve as antennas for light. When they absorb light, they pass its energy (exciton) to the reaction center. There the energy is used to create a charge separation that initiates electron flow through a series of oxidation-reduction reactions.
 Photosynthesis requires the input of energy in the form of light to create a good electron donor and a good electron acceptor.
Photosynthesis requires the input of energy in the form of light to create a good electron donor and a good electron acceptor.  Electrons flow from the electron donor through a series of membrane-bound carriers, including cytochromes, quinones, and iron-sulfur proteins, while protons are pumped across a membrane to create an electrochemical potential. Electron transfer and proton pumping are catalyzed by a membrane complex that is homologous in structure and function to Complex III of mitochondria. The electrochemical potential so produced is the driving force for ATP synthesis from ADP and , catalyzed by a membrane-bound ATP synthase complex closely similar to that of mitochondria and bacteria.
Electrons flow from the electron donor through a series of membrane-bound carriers, including cytochromes, quinones, and iron-sulfur proteins, while protons are pumped across a membrane to create an electrochemical potential. Electron transfer and proton pumping are catalyzed by a membrane complex that is homologous in structure and function to Complex III of mitochondria. The electrochemical potential so produced is the driving force for ATP synthesis from ADP and , catalyzed by a membrane-bound ATP synthase complex closely similar to that of mitochondria and bacteria.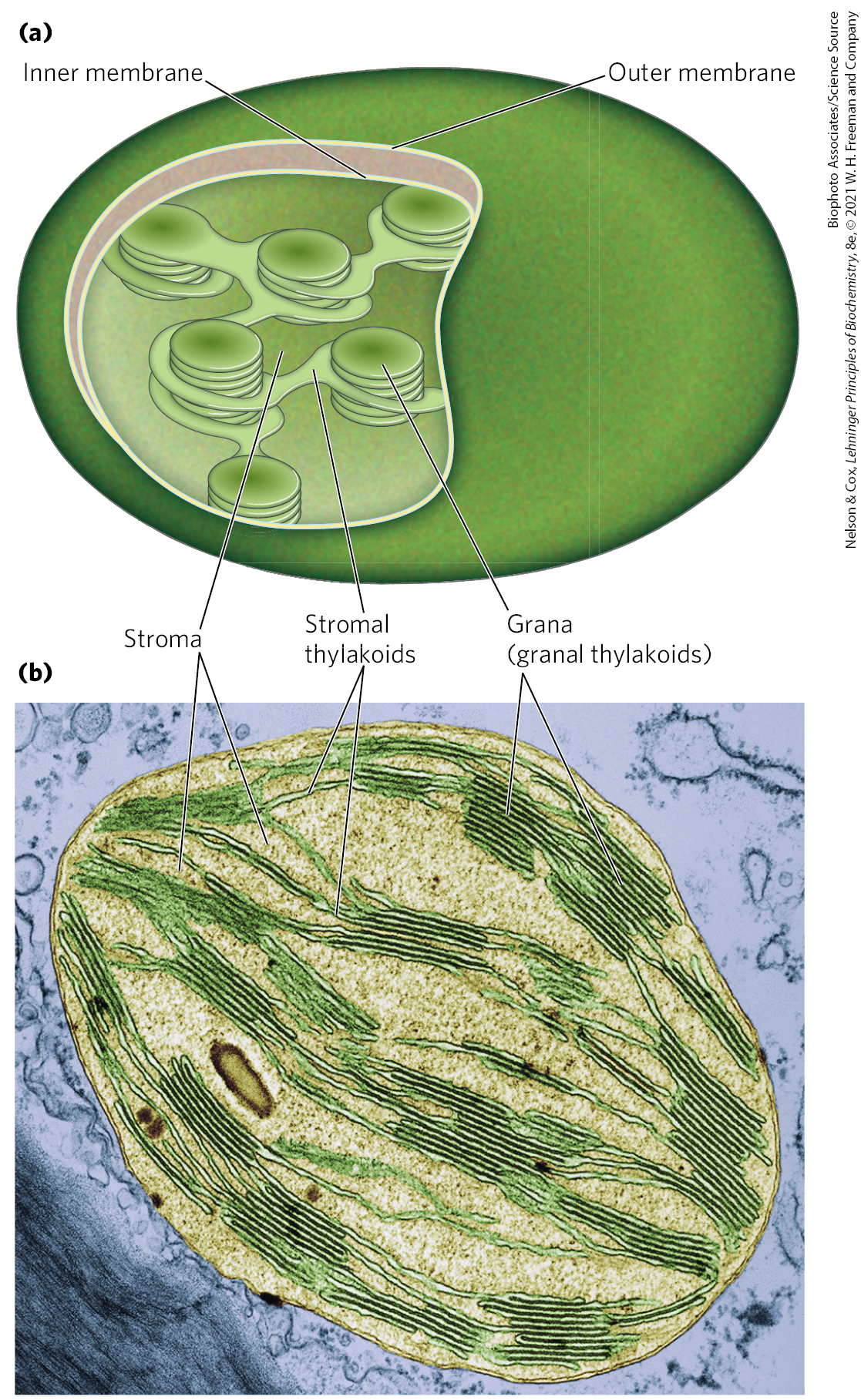
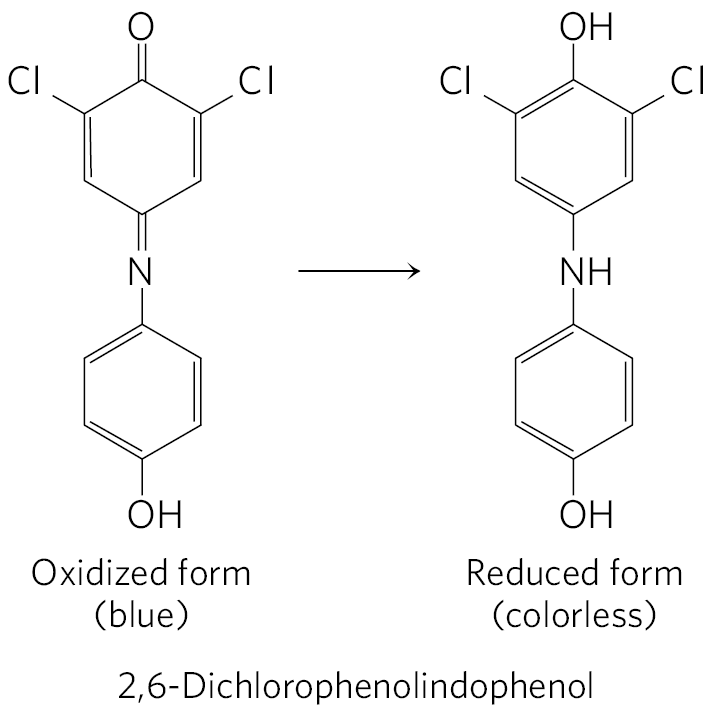
 This was the first evidence that absorbed light energy causes electrons to flow from some electron donor (now known to be ) to an electron acceptor. Moreover, Hill found that was neither required nor reduced to a stable form under these conditions; production could be dissociated from reduction. Several years later, Severo Ochoa showed that is the biological electron acceptor in chloroplasts, according to the equation
This was the first evidence that absorbed light energy causes electrons to flow from some electron donor (now known to be ) to an electron acceptor. Moreover, Hill found that was neither required nor reduced to a stable form under these conditions; production could be dissociated from reduction. Several years later, Severo Ochoa showed that is the biological electron acceptor in chloroplasts, according to the equation

 ), very little fluorescence is observed. Instead, the excited antenna chlorophyll transfers energy directly to a neighboring chlorophyll molecule, which becomes excited as the first molecule returns to its ground state (step
), very little fluorescence is observed. Instead, the excited antenna chlorophyll transfers energy directly to a neighboring chlorophyll molecule, which becomes excited as the first molecule returns to its ground state (step  ). This transfer of energy, exciton transfer, extends to a third, fourth, or subsequent neighbor, until one of a “special pair” of chlorophyll a molecules at the photochemical reaction center is excited (step
). This transfer of energy, exciton transfer, extends to a third, fourth, or subsequent neighbor, until one of a “special pair” of chlorophyll a molecules at the photochemical reaction center is excited (step  ). The special pair of chlorophyll molecules, often designated , are held close enough to each other to share bonding orbitals, and to react as a single compound when excited. In this excited chlorophyll pair, an electron is promoted to a higher-energy orbital. This electron then passes to a nearby electron acceptor that is part of the photosynthetic electron-transfer chain, leaving the reaction-center chlorophyll pair with a missing electron (an “electron hole,” denoted by in
). The special pair of chlorophyll molecules, often designated , are held close enough to each other to share bonding orbitals, and to react as a single compound when excited. In this excited chlorophyll pair, an electron is promoted to a higher-energy orbital. This electron then passes to a nearby electron acceptor that is part of the photosynthetic electron-transfer chain, leaving the reaction-center chlorophyll pair with a missing electron (an “electron hole,” denoted by in  ). The electron acceptor acquires a negative charge in this transaction. The electron lost by the reaction-center chlorophyll pair is replaced by an electron from a neighboring electron-donor molecule (step
). The electron acceptor acquires a negative charge in this transaction. The electron lost by the reaction-center chlorophyll pair is replaced by an electron from a neighboring electron-donor molecule (step  ), which thereby becomes positively charged.
), which thereby becomes positively charged. 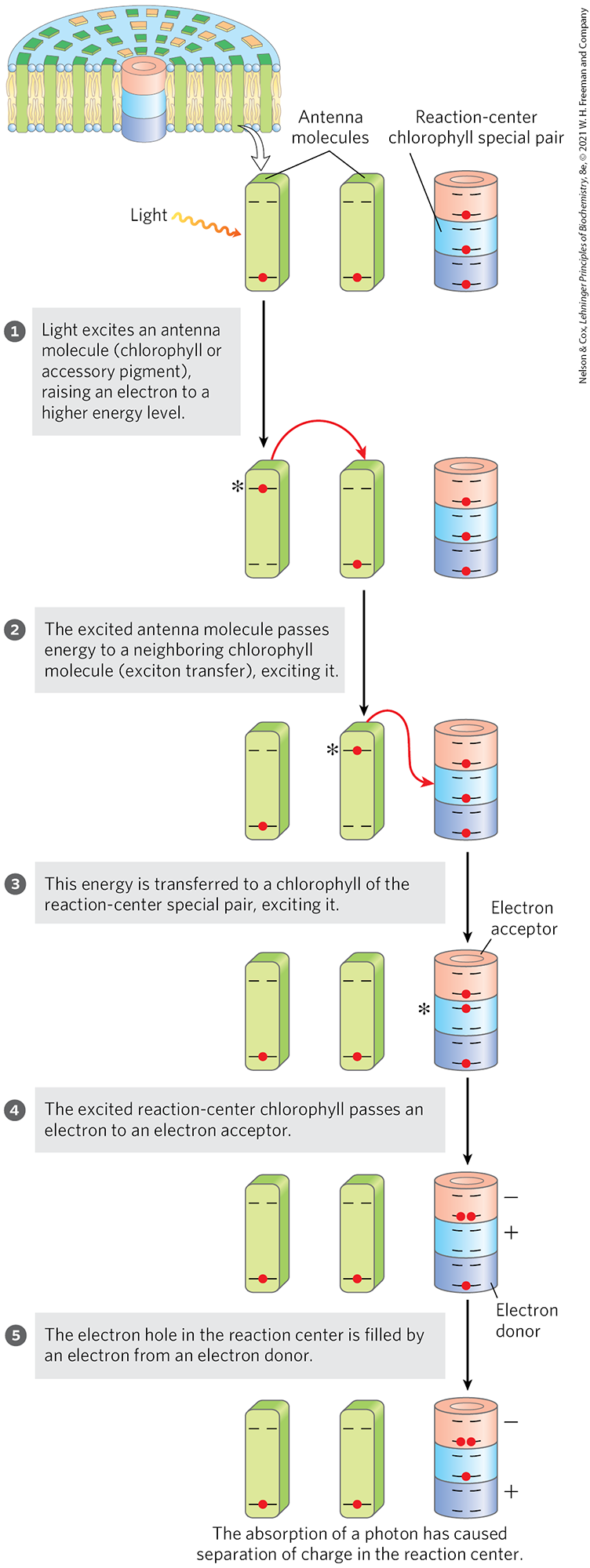
 Photosynthesis takes place in plant chloroplasts, structures enclosed in double membranes and filled with an elaborate system of thylakoid membranes containing the photosynthetic machinery.
Photosynthesis takes place in plant chloroplasts, structures enclosed in double membranes and filled with an elaborate system of thylakoid membranes containing the photosynthetic machinery.