Chapter Review
KEY TERMS
Terms in bold are defined in the glossary.
- signal transduction
- specificity
- sensitivity
- amplification
- enzyme cascade
- modularity
- scaffold proteins
- desensitization
- integration
- divergence
- response localization
- G protein–coupled receptors (GPCRs)
- guanosine nucleotide–binding proteins
- G proteins
- second messenger
- agonist
- antagonist
- β-adrenergic receptors
- seven-transmembrane (7tm) receptors
- guanosine nucleotide–exchange factor (GEF)
- stimulatory G protein ()
- adenylyl cyclase
- cAMP-dependent protein kinase (protein kinase A; PKA)
- consensus sequence
- green fluorescent protein (GFP)
- fluorescence resonance energy transfer (FRET)
- β-adrenergic receptor kinase (βARK)
- β-arrestin (βarr)
- G protein–coupled receptor kinases (GRKs)
- cAMP response element binding protein (CREB)
- inhibitory G protein ()
- adaptor proteins
- AKAPs (A kinase anchoring proteins)
- Ras
- guanosine ,-cyclic monophosphate (cyclic GMP; cGMP)
- cGMP-dependent protein kinase (protein kinase G; PKG)
- GTPase activator protein (GAP)
- regulator of G-protein signaling (RGS)
- NO synthase
- phospholipase C (PLC)
- inositol 1,4,5-trisphosphate
- channel
- protein kinase C (PKC)
- calmodulin (CaM)
- calmodulin-dependent protein kinases (CaM kinases)
- rhodopsin
- rhodopsin kinase
- receptor potential
- receptor Tyr kinase (RTK)
- autophosphorylation
- SH2 domain
- MAPKs
- voltage-gated ion channels
- ionotropic
- metabotropic
- hormone response element (HRE)
- cyclin
- cyclin-dependent protein kinase (CDK)
- ubiquitin
- proteasome
- growth factors
- retinoblastoma protein (pRb)
- oncogene
- proto-oncogene
- tumor suppressor gene
- programmed cell death
- apoptosis
Problems
1. Hormone Experiments in Cell-Free Systems In the 1950s, Earl W. Sutherland, Jr., and his colleagues carried out pioneering experiments to elucidate the mechanism of action of epinephrine and glucagon. Given what you have learned in this chapter about hormone action, interpret each of the experiments described below. Identify substance X and indicate the significance of the results.
- Addition of epinephrine to a homogenate of normal liver resulted in an increase in the activity of glycogen phosphorylase. However, when the homogenate was first centrifuged at a high speed and epinephrine or glucagon was added to the clear supernatant fraction that contains phosphorylase, no increase in the phosphorylase activity occurred.
- When the particulate fraction from the centrifugation in (a) was treated with epinephrine, substance X was produced. The substance was isolated and purified. Unlike epinephrine, substance X activated glycogen phosphorylase when added to the clear supernatant fraction of the centrifuged homogenate.
- Substance X was heat-stable; that is, heat treatment did not affect its capacity to activate phosphorylase. (Hint: Would this be the case if substance X were a protein?) Substance X was nearly identical to a compound obtained when pure ATP was treated with barium hydroxide. (Fig. 8-6 will be helpful.)
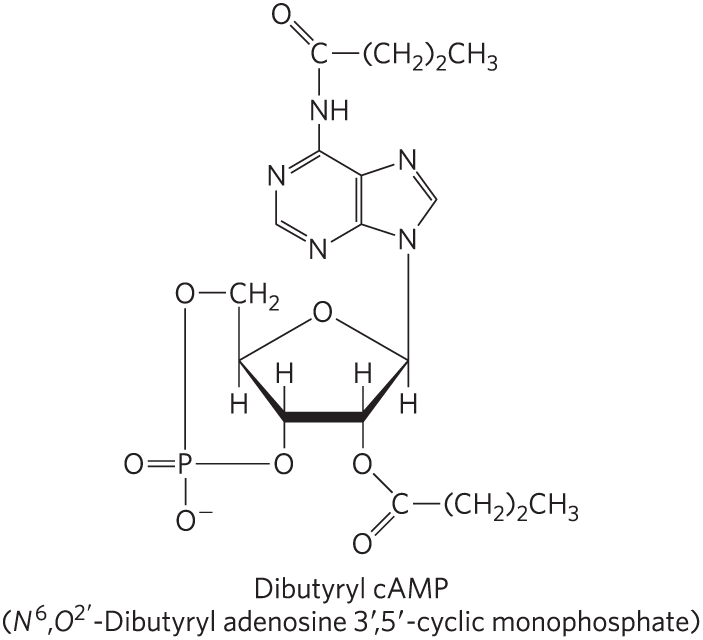
2. Effect of Dibutyryl cAMP versus cAMP on Intact Cells In principle, the physiological effects of epinephrine should be mimicked by addition of cAMP to the target cells. In practice, addition of cAMP to intact target cells elicits only a minimal physiological response. Why? When the structurally related derivative dibutyryl cAMP (shown) is added to intact cells, the expected physiological response is readily apparent. Explain the basis for the difference in cellular response to these two substances. Dibutyryl cAMP is widely used in studies of cAMP function.
3. Effect of Cholera Toxin on Adenylyl Cyclase The gram-negative bacterium Vibrio cholerae produces a protein, cholera toxin ( 90,000), that is responsible for the characteristic symptoms of cholera: extensive loss of body water and through continuous, debilitating diarrhea. If body fluids and are not replaced, severe dehydration results; untreated, the disease is often fatal. When the cholera toxin gains access to the human intestinal tract, it binds tightly to specific sites in the plasma membrane of the epithelial cells lining the small intestine, causing adenylyl cyclase to undergo prolonged activation (hours or days).
- What is the expected effect of cholera toxin on [cAMP] in the intestinal cells?
- Based on the information above, suggest how cAMP normally functions in intestinal epithelial cells.
- Suggest a possible treatment for cholera.
4. Mutations in PKA Explain how mutations in the R or C subunit of cAMP-dependent protein kinase (PKA) might lead to (a) a constantly active PKA or (b) a constantly inactive PKA.
5. Therapeutic Effects of Albuterol The respiratory symptoms of asthma result from constriction of the bronchi and bronchioles of the lungs, caused by contraction of the smooth muscle of their walls. Raising [cAMP] in the smooth muscle reverses the constriction of the bronchi and bronchioles. Explain the therapeutic effects of albuterol, an inhaled β-adrenergic agonist, in treating asthma. Would you expect this drug to have any side effects? If so, what design change could you make to the drug to minimize side effects?
6. Termination of Hormonal Signals Signals carried by hormones must eventually be terminated. Describe several mechanisms for signal termination.
7. Using FRET to Explore Protein-Protein Interactions In Vivo Figure 12-9 shows the interaction between β-arrestin and the β-adrenergic receptor. How would you use FRET (see Box 12-1) to demonstrate this interaction in living cells? Which proteins would you fuse? Which wavelengths would you use to illuminate the cells, and which wavelengths would you monitor? What would you expect to observe if the interaction occurred? If it did not occur? How might you explain the failure of this approach to demonstrate this interaction?
8. EGTA Injection EGTA (ethylene glycol-bis(β-amino ethyl ether)--tetraacetic acid) is a chelating agent with high affinity and specificity for . By microinjecting a cell with an appropriate -EGTA solution, an experimenter can prevent cytosolic from rising above . How would EGTA microinjection affect a cell’s response to vasopressin (see Table 12-4)? To glucagon?
9. Amplification and Termination of Hormonal Signals In the β-adrenergic system, which of these contributes to the amplification of the signal (epinephrine) and which to the termination of the signal? Do any contribute to both amplification and termination of the signal?
- One activates many adenylyl cyclase molecules.
- One protein kinase A (PKA) phosphorylates many target proteins.
- The intrinsic GTPase of G protein converts bound GTP to GDP.
- A phosphodiesterase acts on many molecules of cAMP.
- One epinephrine molecule activates many adrenergic receptors.
- One protein kinase phosphorylates many molecules of another protein kinase.
10. The Insulin Signaling System Place these components of the insulin receptor system in the order in which they occur in the sequence of events triggered by insulin: MEK, Ras, ERK, GRK, Raf, Sos, IRS1, PKA, Grb2. Some of these may not participate in that path.
11. Mutations in ras How would a mutation in ras that leads to formation of a Ras protein with no GTPase activity affect a cell’s response to insulin?
12. Differences among G Proteins Compare the G protein , which acts in transducing the signal from β-adrenergic receptors, and the G protein Ras. What properties do they share? How do they differ? What is the functional difference between and ?
13. Mechanisms for Regulating Protein Kinases Identify eight general types of protein kinases found in eukaryotic cells, and explain what factor is directly responsible for activating each type.
14. Nonhydrolyzable GTP Analogs Many enzymes can hydrolyze GTP between the β and γ phosphates. The GTP analog β,γ-imidoguanosine -triphosphate (Gpp(NH)p), shown here, cannot be hydrolyzed between the β and γ phosphates.
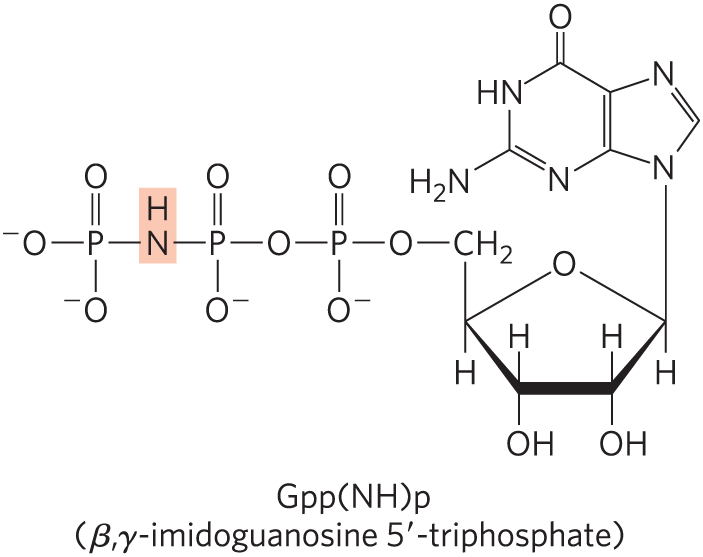
Predict the effect of microinjection of Gpp(NH)p into a myocyte on the cell’s response to β-adrenergic stimulation.
15. Visual Desensitization Oguchi disease is an inherited form of night blindness. Affected individuals are slow to recover vision after a flash of bright light against a dark background, such as the headlights of a car on the freeway. Suggest what the molecular defect(s) might be in Oguchi disease. Explain in molecular terms how this defect would account for night blindness.
16. Effect of a Permeant cGMP Analog on Rod Cells An analog of cGMP, 8-Br-cGMP, will permeate cellular membranes, is only slowly degraded by a rod cell’s PDE activity, and is as effective as cGMP in opening the gated channel in the cell’s outer segment. If you suspended rod cells in a buffer containing a relatively high [8-Br-cGMP], then illuminated the cells while measuring their membrane potential, what would you expect to see?
17. Effect of Insulin on Glycogen Synthesis Protein kinase B (PKB) inactivates glycogen synthase kinase (GSK3), and GSK3 inactivates glycogen synthase. Predict the effect of insulin on glycogen synthesis.
18. Role of Intrinsically Disordered Regions of Signaling Proteins Signaling proteins, including protein kinases, often have intrinsically disordered regions (IDRs) that are important in signaling. Describe a case in which IDRs and their interactions with other proteins are important in signaling.
19. The Action Potential Place these events in the order in which they occur after a presynaptic neuron releases acetylcholine into the synaptic cleft.
- Vesicles containing a neurotransmitter fuse with the cell membrane.
- Ligand-gated channels open, causing an influx of ions.
- Voltage-gated channels open in the axon.
- Membrane depolarization triggers voltage-gated channels to open.
- Local membrane depolarization in the axon triggers an efflux of .
20. Hot and Cool Taste Sensations The sensations of heat and cold are transduced by a group of temperature-gated cation channels. For example, TRPV1, TRPV3, and TRPM8 are usually closed, but they open at different temperatures. TRPV1 opens at , TRPV3 opens at , and TRPM8 opens at . These channel proteins are expressed in sensory neurons known to be responsible for temperature sensation.
- Propose a reasonable model to explain how exposing a sensory neuron containing TRPV1 to high temperature leads to a sensation of heat.
- Capsaicin, one of the active ingredients in “hot” peppers, is an agonist of TRPV1. Capsaicin shows 50% activation of the TRPV1 response at a concentration of 32 nm — a property known as . Explain why even a very few drops of hot pepper sauce can taste very “hot” without actually burning you.
- Menthol, one of the active ingredients in mint, is an agonist of TRPM8 and TRPV3 . What sensation would you expect from contact with low levels of menthol? With high levels?
21. Oncogenes, Tumor Suppressor Genes, and Tumors For each of the situations listed, provide a plausible explanation for how it could lead to unrestricted cell division.
- Colon cancer cells often contain mutations in the gene encoding the prostaglandin receptor. is a growth factor required for the division of cells in the gastrointestinal tract.
- Kaposi sarcoma, a common tumor in people with untreated AIDS, is caused by a virus carrying a gene for a protein similar to the chemokine receptors CXCR1 and CXCR2. Chemokines are cell-specific growth factors.
- Adenovirus, a tumor virus, carries a gene for the protein E1A, which binds to the retinoblastoma protein, pRb. (Hint: See Fig. 12-40.)
- An important feature of many oncogenes and tumor suppressor genes is their cell-type specificity. For example, mutations in the receptor are not typically found in lung tumors. Explain this observation. (Note that acts through a GPCR in the plasma membrane.)
22. Mutations in Tumor Suppressor Genes and Oncogenes Explain why mutations in tumor suppressor genes are recessive (both copies of the gene must be defective for the regulation of cell division to be defective), whereas mutations in oncogenes are dominant.
23. Retinoblastoma in Children Explain why some children with retinoblastoma develop multiple tumors of the retina in both eyes, whereas others have a single tumor in only one eye.
24. Specificity of a Signal for a Single Cell Type Discuss the validity of the proposition that a signaling molecule (hormone, growth factor, or neurotransmitter) elicits identical responses in different types of target cells if those cells contain identical receptors.
 Effect of Cholera Toxin on Adenylyl Cyclase The gram-negative bacterium Vibrio cholerae produces a protein, cholera toxin ( 90,000), that is responsible for the characteristic symptoms of cholera: extensive loss of body water and through continuous, debilitating diarrhea. If body fluids and are not replaced, severe dehydration results; untreated, the disease is often fatal. When the cholera toxin gains access to the human intestinal tract, it binds tightly to specific sites in the plasma membrane of the epithelial cells lining the small intestine, causing adenylyl cyclase to undergo prolonged activation (hours or days).
Effect of Cholera Toxin on Adenylyl Cyclase The gram-negative bacterium Vibrio cholerae produces a protein, cholera toxin ( 90,000), that is responsible for the characteristic symptoms of cholera: extensive loss of body water and through continuous, debilitating diarrhea. If body fluids and are not replaced, severe dehydration results; untreated, the disease is often fatal. When the cholera toxin gains access to the human intestinal tract, it binds tightly to specific sites in the plasma membrane of the epithelial cells lining the small intestine, causing adenylyl cyclase to undergo prolonged activation (hours or days).DATA ANALYSIS PROBLEM
25. Exploring Taste Sensation in Mice Pleasing tastes are an evolutionary adaptation to encourage animals to consume nutritious foods. Zhao and coauthors (2003) examined the two major pleasurable taste sensations: sweet and umami. Umami is a “distinct savory taste” triggered by amino acids, especially aspartate and glutamate, and it probably encourages animals to consume protein-rich foods. Monosodium glutamate (MSG) is a flavor-enhancer that exploits this sensitivity.
At the time the article was published, three taste receptor proteins for sweet and umami had been tentatively characterized: T1R1, T1R2, and T1R3. These proteins function as heterodimeric receptor complexes: T1R1-T1R3 was tentatively identified as the umami receptor, and T1R2-T1R3 as the sweet receptor. It was not clear how taste sensation was encoded and sent to the brain, and two possible models had been suggested. In the cell-based model, individual taste-sensing cells express only one kind of receptor; that is, there are “sweet cells,” “bitter cells,” “umami cells,” and so on, and each type of cell sends its information to the brain via a different nerve. The brain “knows” which taste is detected by the identity of the nerve fiber that transmits the message. In the receptor-based model, individual taste-sensing cells have several kinds of receptors and send different messages along the same nerve fiber to the brain, the message depending on which receptor is activated. Also unclear at the time was whether there was any interaction between the different taste sensations, or whether parts of one taste-sensing system were required for other taste sensations.
Previous work had shown that different taste receptor proteins are expressed in nonoverlapping sets of taste receptor cells. Which model does this support? Explain your reasoning.
Zhao and colleagues constructed a set of “knockout mice” — mice homozygous for loss-of-function alleles for one of the three receptor proteins, T1R1, T1R2, or T1R3 — and double-knockout mice with nonfunctioning T1R2 and T1R3. The researchers measured the taste perception of these mice by measuring their “lick rate” of solutions containing different taste molecules. Mice will lick the spout of a feeding bottle with a pleasant-tasting solution more often than one with an unpleasant-tasting solution. The researchers measured relative lick rates: how often the mice licked a sample solution compared with water. A relative lick rate of 1 indicated no preference; , an aversion; and , a preference.
All four types of knockout strains had the same responses to salt and bitter tastes as did wild-type mice. Which of the above issues did this experiment address? What do you conclude from these results?
The researchers then studied umami taste reception by measuring the relative lick rates of the different mouse strains with different quantities of MSG in the feeding solution. Note that the solutions also contained inosine monophosphate (IMP), a strong potentiator of umami taste reception (and a common ingredient in ramen soups, along with MSG), and amiloride, which suppresses the pleasant salty taste imparted by the sodium of MSG. The results are shown in the graph.
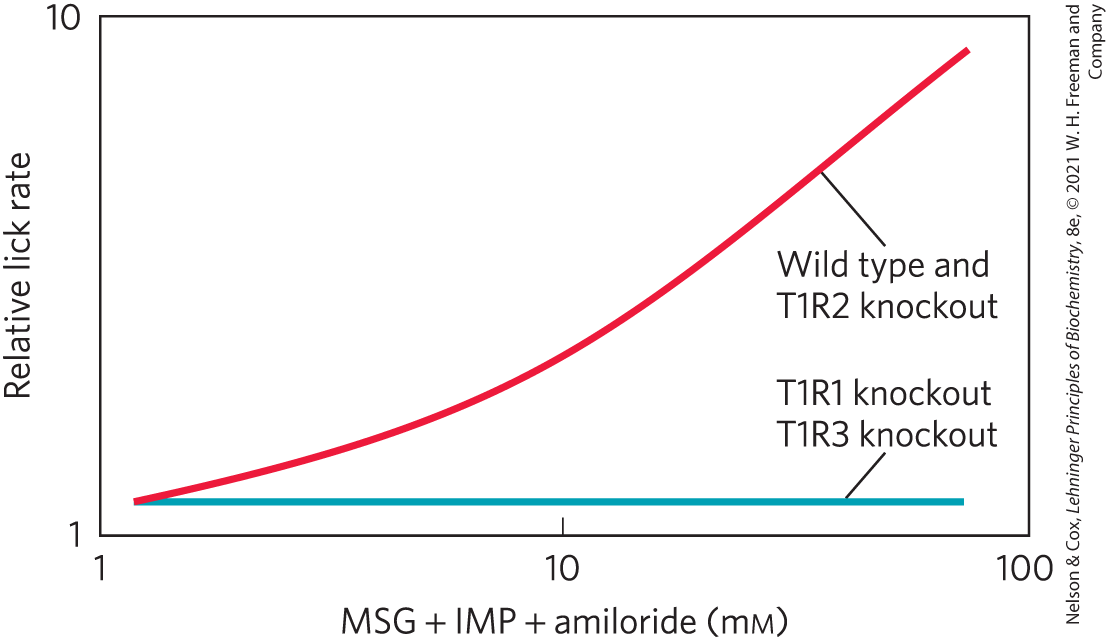
Are these data consistent with the umami taste receptor consisting of a heterodimer of T1R1 and T1R3? Why or why not?
Which model(s) of taste encoding does this result support? Explain your reasoning.
Zhao and coworkers then performed a series of similar experiments using sucrose as a sweet taste. These results are shown below.
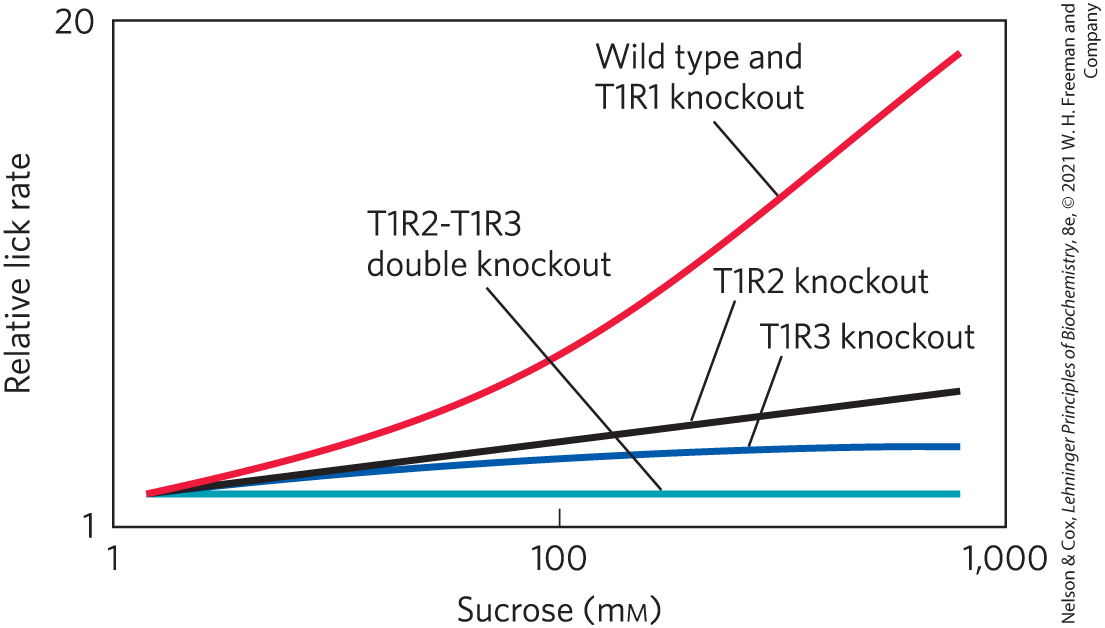
Are these data consistent with the sweet taste receptor consisting of a heterodimer of T1R2 and T1R3? Why or why not?
There were some unexpected responses at very high sucrose concentrations. How do these complicate the idea of a heterodimeric system as presented above?
In addition to sugars, humans also taste other compounds (e.g., saccharin and the peptides monellin and aspartame) as sweet; mice do not taste these as sweet. Zhao and coworkers inserted into TIR2-knockout mice a copy of the human T1R2 gene under the control of the mouse T1R2 promoter. These modified mice now tasted monellin and saccharin as sweet. The researchers then went further, adding to T1R1-knockout mice the RASSL protein — a G protein–linked receptor for the synthetic opiate spiradoline; the RASSL gene was under the control of a promoter that could be induced by feeding the mice tetracycline. These mice did not prefer spiradoline in the absence of tetracycline; in the presence of tetracycline, they showed a strong preference for nanomolar concentrations of spiradoline.
Do these results strengthen your conclusions about the mechanism of taste sensation?
Reference
- Zhao, G.Q., Y. Zhang, M.A. Hoon, J. Chandrashekar, I. Erlenbach, N.J.P. Ryba, and C. Zuker. 2003. The receptors for mammalian sweet and umami taste. Cell 115:255–266.