Chapter Review
Key Terms
Terms in bold are defined in the glossary.
- chromosome
- phenotype
- mutation
- gene
- regulatory sequence
- plasmid
- intron
- exon
- simple-sequence DNA
- satellite DNA
- centromere
- telomere
- supercoil
- relaxed DNA
- topology
- linking number
- specific linking difference
- superhelical density (σ)
- topoisomers
- topoisomerases
- catenane
- plectonemic
- solenoidal
- chromatin
- histones
- nucleosome
- histone exchange factors
- heterochromatin
- topologically associating domains (TADs)
- long noncoding RNA (lncRNA)
- epigenetic
- euchromatin
- chromosome territory
- Barr body
- SMC proteins
- cohesins
- condensins
- nucleoid
Problems
1. Packaging of DNA in a Virus Bacteriophage T2 has a DNA of molecular weight contained in a head about 210 nm long. Calculate the length of the DNA and compare it with the length of the T2 head. Assume the molecular weight of a nucleotide base pair is 650 and that the DNA is in the B form and relaxed.
2. The DNA of Phage M13 The base composition of the DNA within a bacteriophage M13 viral particle is A, 23%; T, 36%; G, 21%; C, 20%. What does this tell you about this DNA molecule?
3. The Mycoplasma Genome The complete genome of the simplest bacterium known, Mycoplasma genitalium, is a circular DNA molecule with 580,070 bp. Calculate the molecular weight and contour length (when relaxed) of this molecule. What is for the Mycoplasma chromosome? If , what is Lk?
4. Size of Eukaryotic Genes An enzyme isolated from rat liver has 192 amino acid residues and is encoded by a gene with 1,440 bp. Explain the relationship between the number of amino acid residues in the enzyme and the number of nucleotide pairs in its gene.
5. Linking Number A closed-circular DNA molecule in its relaxed form has an Lk of 500. Approximately how many base pairs are in this DNA? How is Lk altered (increases, decreases, doesn’t change, becomes undefined) when
- a protein complex binds to form a nucleosome,
- one DNA strand is broken,
- DNA gyrase and ATP are added to the DNA solution, or
- the double helix is denatured by heat?
6. DNA Topology In the presence of a eukaryotic condensin and a bacterial type II topoisomerase, the Lk of a relaxed closed-circular DNA molecule does not change. However, the DNA becomes highly knotted.
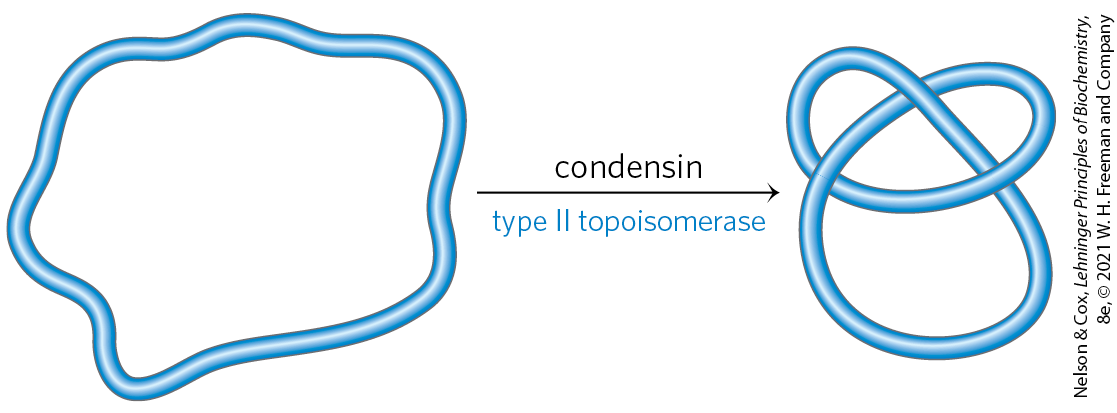
Formation of the knots requires breakage of the DNA, passage of a segment of DNA through the break, and religation by the topoisomerase. Given that every reaction of the topoisomerase would be expected to result in a change in linking number, how can Lk remain the same?
7. Superhelical Density Bacteriophage λ infects E. coli by integrating its DNA into the bacterial chromosome. The success of this recombination depends on the topology of the E. coli DNA. When the superhelical density (σ) of the E. coli DNA is greater than −0.045, the probability of integration is less than 20%; when σ is less than −0.06, the probability is >70%. Plasmid DNA isolated from an E. coli culture is found to have a length of 13,800 bp and an Lk of 1,222. Calculate σ for this DNA and predict the likelihood that bacteriophage λ will be able to infect this culture.
8. Altering Linking Number
- What is the Lk of a 5,000 bp circular duplex DNA molecule with a nick in one strand?
- What is the Lk of the molecule in (a) when the nick is sealed (relaxed)?
- How would the Lk of the molecule in (b) be affected by the action of a single molecule of E. coli topoisomerase I?
- What is the Lk of the molecule in (b) after eight enzymatic turnovers by a single molecule of DNA gyrase in the presence of ATP?
- What is the Lk of the molecule in (d) after four enzymatic turnovers by a single molecule of bacterial type I topoisomerase?
- What is the Lk of the molecule in (d) after binding of one nucleosome core?
9. Chromatin The agarose gel shown, in which the thick bands represent DNA, helped researchers define nucleosome structure. They generated this result by briefly treating chromatin with an enzyme that degrades DNA, then removing all protein and subjecting the purified DNA to electrophoresis. Numbers at the side of the gel denote the position to which a linear DNA of the indicated size would migrate. What does this gel demonstrate about chromatin structure? Why are the DNA bands thick and spread out rather than sharply defined?

10. DNA Structure Explain how the underwinding of a B-DNA helix might facilitate or stabilize the formation of Z-DNA (see Fig. 8-17).
11. Maintaining DNA Structure
- Describe two structural features required for a DNA molecule to maintain a negatively supercoiled state.
- List three structural changes that become more favorable when a DNA molecule is negatively supercoiled.
- What E. coli enzyme, with the aid of ATP, can generate negative superhelicity in DNA?
- Describe the physical mechanism by which this enzyme acts.
12. Yeast Artificial Chromosomes (YACs) Researchers use YACs to clone large pieces of DNA in yeast cells. What three types of DNA sequence do researchers require to ensure proper replication and propagation of a YAC in a yeast cell?
13. Nucleoid Structure in Bacteria In bacteria, DNA topology affects the transcription of a subset of genes, with expression increasing or (more often) decreasing when the DNA is relaxed. Following cleavage of a bacterial chromosome at a specific site by a restriction enzyme (one that cuts at a long, and thus rare, sequence), only nearby genes (within 10,000 bp) exhibit either an increase or a decrease in expression. The transcription of genes elsewhere in the chromosome is unaffected. Explain. (Hint: See Fig. 24-35.)
14. DNA Topology When DNA is subjected to electrophoresis in an agarose gel, shorter molecules migrate faster than longer ones. Closed-circular DNAs of the same size but with different linking numbers also can be separated on an agarose gel: topoisomers that are more supercoiled, and thus more condensed, migrate faster through the gel. In the gel shown, purified plasmid DNA has migrated from top to bottom. There are two bands, with the faster band much more prominent.

- What are the DNA species in the two bands?
- If topoisomerase I were added to a solution of this DNA, what would happen to the upper and lower bands after electrophoresis?
- If DNA ligase were added to the DNA, would the appearance of the bands change? Explain your answer.
- If DNA gyrase plus ATP were added to the DNA after the addition of DNA ligase, how would the band pattern change?
15. DNA Topoisomers When DNA is subjected to electrophoresis in an agarose gel, shorter molecules migrate faster than longer ones. Closed-circular DNAs of the same size but different linking number also can be separated on an agarose gel: topoisomers that are more supercoiled, and thus more condensed, migrate faster through the gel — from top to bottom in the two gels shown. An investigator added a dye, chloroquine, to these gels. Chloroquine intercalates between base pairs and stabilizes a more underwound DNA structure. When the dye binds to a relaxed closed-circular DNA, the DNA is underwound where the dye binds, and unbound regions take on positive supercoils to compensate. In the experiment shown here, an investigator used topoisomerases to make preparations of the same DNA circle with different superhelical densities (σ). Completely relaxed DNA migrated to the position labeled N (for nicked), and highly supercoiled DNA (above the limit where individual topoisomers can be distinguished) migrated to the position labeled X.
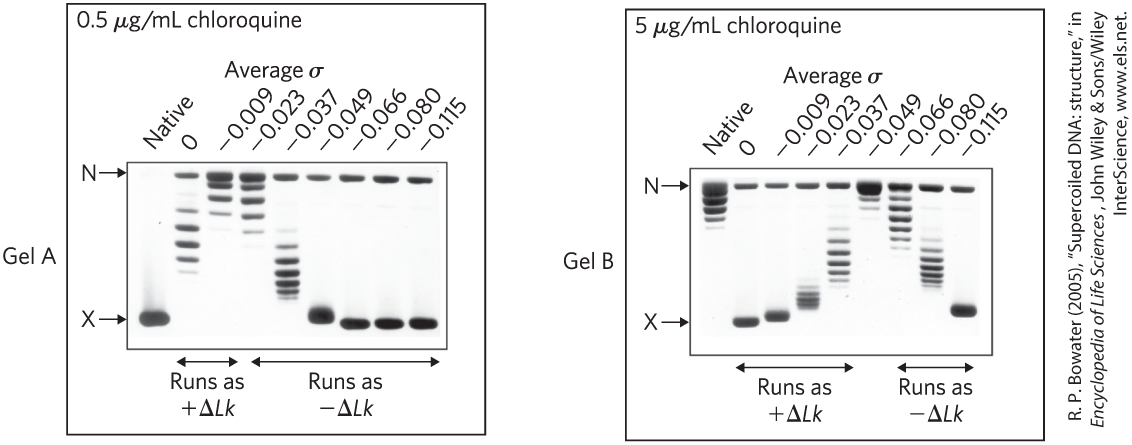
- In gel A, why does the lane (i.e., DNA prepared so that , on average) have multiple bands?
- In gel B, is the DNA from the preparation positively or negatively supercoiled in the presence of the intercalating dye?
- In both gels, the lane has two bands, one a highly supercoiled DNA and one relaxed. Propose a reason for the presence of relaxed DNA in these lanes (and others).
- The native DNA (leftmost lane in each gel) is the same DNA circle isolated from bacterial cells and untreated. What is the approximate superhelical density of this native DNA?
16. Nucleosomes The human genome comprises just over 3.1 billion base pairs. Assuming it contains nucleosomes that are spaced as described in this chapter, how many molecules of histone H2A are present in one somatic human cell? (Ignore reductions in H2A due to its replacement in some regions by H2A variants.) How would the number change after DNA replication but before cell division?
17. Bacterial DNA Topoisomerase IV The gene encoding topoisomerase IV in E. coli is essential, even though another type II topoisomerase (topoisomerase II or gyrase) is present. Suggest a reason for the requirement for topoisomerase IV.
18. Chromosome Topology Eukaryotic chromosomes are linear DNA molecules, yet the DNA of a chromosome retains a high level of underwinding (supercoiling) throughout its length. How does the organization of chromosomal DNA into loops called TADs contribute to the maintenance of supercoiling?
Data Analysis Problem
19. Defining the Functional Elements of Yeast Chromosomes Figure 24-7 shows the major structural elements of a chromosome of budding yeast (S. cerevisiae). Heiter, Mann, Snyder, and Davis (1985) determined the properties of some of these elements. They based their study on the finding that in yeast cells, plasmids (which have genes and an origin of replication) act differently from chromosomes (which have these elements plus centromeres and telomeres) during mitosis. The plasmids are not manipulated by the mitotic apparatus and segregate randomly between daughter cells. Without a selectable marker to force the host cells to retain them (see Fig. 9-4), these plasmids are rapidly lost. In contrast, chromosomes, even without a selectable marker, are manipulated by the mitotic apparatus and are lost at a very low frequency (about per cell division).
Heiter and colleagues set out to determine the important components of yeast chromosomes by constructing plasmids with various parts of chromosomes and observing whether these “synthetic chromosomes” segregated properly during mitosis. To measure the frequencies of different types of failed chromosome segregation, the researchers needed a rapid assay to determine the number of copies of synthetic chromosomes present in different cells. The assay took advantage of the fact that wild-type yeast colonies are white whereas certain adenine-requiring mutants yield red colonies on nutrient media; cells lack functional AIR carboxylase (the enzyme of step in Fig. 22-35) and accumulate AIR (5-aminoimidazole ribonucleotide) in their cytoplasm, and the excess AIR is converted to a conspicuous red pigment. The other part of the assay involved the gene SUP11, which encodes an ochre suppressor (a type of nonsense suppressor in which a termination codon specifies an amino acid) that suppresses the phenotype of some mutants.
Heiter and coworkers started with a diploid strain of yeast homozygous for ; these cells are red. When the mutant cells contain one copy of SUP11, the metabolic defect is partly suppressed and the cells are pink. When the cells contain two or more copies of SUP11, the defect is completely suppressed and the cells are white.
The researchers inserted one copy of SUP11 into synthetic chromosomes containing various elements thought to be important in chromosome function, and then observed how well these chromosomes were passed from one generation to the next. These pink cells were plated on nonselective media, and the behavior of the synthetic chromosomes was observed. Heiter and coworkers looked for colonies in which the synthetic chromosomes segregated improperly at the first division after plating, giving rise to a colony that was half one genotype and half the other. Because yeast cells are nonmotile, this would be a sectored colony, with one half one color and the other half another color.
- One way for the mitotic process to fail is nondisjunction: the chromosome replicates but the sister chromatids fail to separate, so both copies of the chromosome end up in the same daughter cell. Explain how nondisjunction of the synthetic chromosome would give rise to a colony that is half red and half white.
- Another way for the mitotic process to fail is chromosome loss: the chromosome does not enter the daughter nucleus or is not replicated. Explain how loss of the synthetic chromosome would give rise to a colony that is half red and half pink.
By counting the frequency of the different colony types, Heiter and colleagues could estimate the frequency of these aberrant mitotic events with different types of synthetic chromosome. First, they explored the requirement for centromeric sequences by constructing synthetic chromosomes with DNA fragments of different sizes containing a known centromere. Their results are shown here.
Synthetic chromosome Size of centromere-containing fragment (kbp) Chromosome loss (%) Nondisjunction (%) 1
None
—
>50
2
0.63
1.6
1.1
3
1.6
1.9
0.4
4
3.0
1.7
0.35
5
6.0
1.6
0.35
- Based on these data, what can you conclude about the size of the centromere required for normal mitotic segregation? Explain your reasoning.
- All the synthetic chromosomes created in these experiments were circular and lacked telomeres. Explain how they could be replicated more-or-less properly.
Heiter and colleagues next constructed a series of linear synthetic chromosomes that included the functional centromeric sequence and telomeres, and they measured the total mitotic error frequency (% loss + % nondisjunction) as a function of size.
Synthetic chromosome Size (kbp) Total error frequency (%) 6
15
11.0
7
55
1.5
8
95
0.44
9
137
0.14
- Based on these data, what can you conclude about the chromosome size required for normal mitotic segregation? Explain your reasoning.
- Normal yeast chromosomes are linear, range from 250 kbp to 2,000 kbp in length, and, as noted above, have a mitotic error frequency of about per cell division. Extrapolating the results from (e), do the centromeric and telomeric sequences used in these experiments explain the mitotic stability of normal yeast chromosomes, or must other elements be involved? Explain your reasoning. (Hint: A plot of log of error frequency vs. length will be helpful.)
 in
in Reference
- Heiter, P., C. Mann, M. Snyder, and R.W. Davis. 1985. Mitotic stability of yeast chromosomes: A colony color assay that measures nondisjunction and chromosome loss. Cell 40:381–392.