Chapter Review
KEY TERMS
Terms in bold are defined in the glossary.
- ligand
- binding site
- induced fit
- hemoglobin
- heme
- porphyrin
- globins
- globin fold
- equilibrium expression
- association constant,
- dissociation constant,
- allosteric protein
- modulator
- homotropic
- heterotropic
- Hill equation
- Hill coefficient
- Bohr effect
- immune response
- lymphocytes
- antibody
- immunoglobulin (Ig)
- B lymphocyte (B cell)
- T lymphocyte (T cell)
- antigen
- epitope
- hapten
- immunoglobulin fold
- polyclonal antibodies
- monoclonal antibodies
- immunoblotting
- Western blotting
- myosin
- actin
- myofibril
- sarcomere
Problems
1. Relationship between Affinity and Dissociation Constant Protein A has a binding site for ligand X with a of . Protein B has a binding site for ligand X with a of . Calculate the for each protein. Which protein has a higher affinity for ligand X? Explain your reasoning.
2. Modeling Apparent Negative Cooperativity Which of these situations would produce a Hill plot with ? Explain your reasoning in each case.
- The protein has multiple subunits, each with a single ligand-binding site. Ligand binding to one site decreases the binding affinity of other sites for the ligand.
- The protein is a single polypeptide with two ligand-binding sites, each having a different affinity for the ligand.
- The protein is a single polypeptide with a single ligand-binding site. As purified, the protein preparation is heterogeneous, containing some protein molecules that are partially denatured and thus have a lower binding affinity for the ligand.
- The protein has multiple subunits, each with a single ligand-binding site. Ligands bind independently to each site, do not affect the binding affinity of other sites, and bind with identical affinities.
3. Reversible Ligand Binding I The protein calcineurin binds to the protein calmodulin with an association rate of and an overall dissociation constant, , of 10 nm. Calculate the dissociation rate, , including appropriate units.
4. Reversible Ligand Binding II The E. coli nickel-binding protein binds to its ligand, , with a of 100 nm. Calculate the concentration when the fraction of binding sites occupied by the ligand (Y) is (a) 0.25, (b) 0.6, (c) 0.95.
5. Reversible Ligand Binding III You are a technician in a biochemistry lab running receptor binding experiments. The target membrane-bound receptor has been partially purified from mouse, rat, and human cell lines. Using various concentrations of the same radioactive ligand for each receptor in a saturation binding assay, you generate the binding data shown in the table. The dependent variable, Y, is the fraction of binding sites occupied by the ligand.
Ligand concentration (nM) Y Mouse receptor Rat receptor Human receptor 0.2
0.048
0.29
0.17
0.5
0.11
0.50
0.33
1.0
0.20
0.67
0.50
4.0
0.50
0.89
0.80
10
0.71
0.95
0.91
20
0.83
0.97
0.95
50
0.93
0.99
0.98
- Determine the mouse receptor in this binding experiment.
- Which receptor binds most tightly to this ligand?
6. Reversible Ligand Binding IV Exposure to carbon monoxide can lead to unconsciousness and ultimately death. Suffocation occurs when hemoglobin is half-saturated with CO — that is, when only two of every four oxygen-binding sites are occupied with CO. Explain why death may occur at this point, even though half of the oxygen-binding sites are still available to transport . (Hint: See Box 5-1.)
7. Cooperativity in Hemoglobin Under appropriate conditions, hemoglobin dissociates into its four subunits. The isolated α subunit binds oxygen, but the -saturation curve is hyperbolic rather than sigmoid. In addition, the binding of oxygen to the isolated α subunit is not affected by the presence of , , or BPG. What do these observations indicate about the source of the cooperativity in hemoglobin?
8. Oxygen Binding to Hemoglobin The solid curve in the plot shown is an -binding curve for human hemoglobin. For each condition, indicate whether the stated physiological change would shift the curve to the left (dashed curve), produce no change (black curve), or shift the curve to the right (dashed curve).
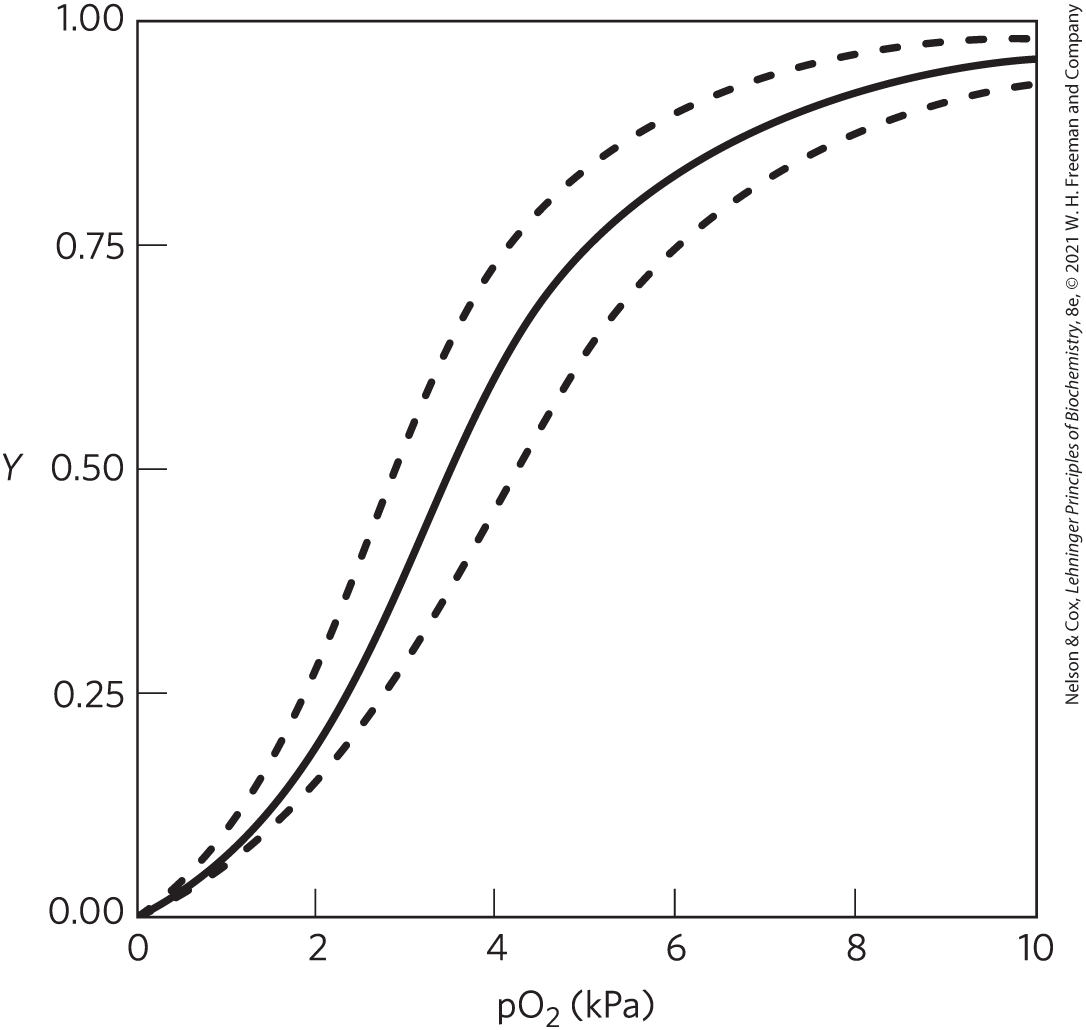
- an increase in the concentration of
- an increase in the proton concentration (decrease in pH)
- an increase in the concentration of 2,3-bisphosphoglycerate (BPG)
9. Comparison of Fetal and Maternal Hemoglobins Studies of oxygen transport in pregnant mammals show that the -saturation curves of fetal and maternal blood are markedly different when measured under the same conditions. Fetal erythrocytes contain a structural variant of hemoglobin, HbF, consisting of two α and two γ subunits (), whereas maternal erythrocytes contain HbA ().
- Which hemoglobin has a higher affinity for oxygen under physiologic conditions?
- What is the physiological significance of the different affinities?
When all the BPG is carefully removed from samples of HbA and HbF, the measured -saturation curves (and consequently the affinities) are displaced to the left. However, HbA now has a greater affinity for oxygen than does HbF. When BPG is reintroduced, the -saturation curves return to normal, as shown in the graph.
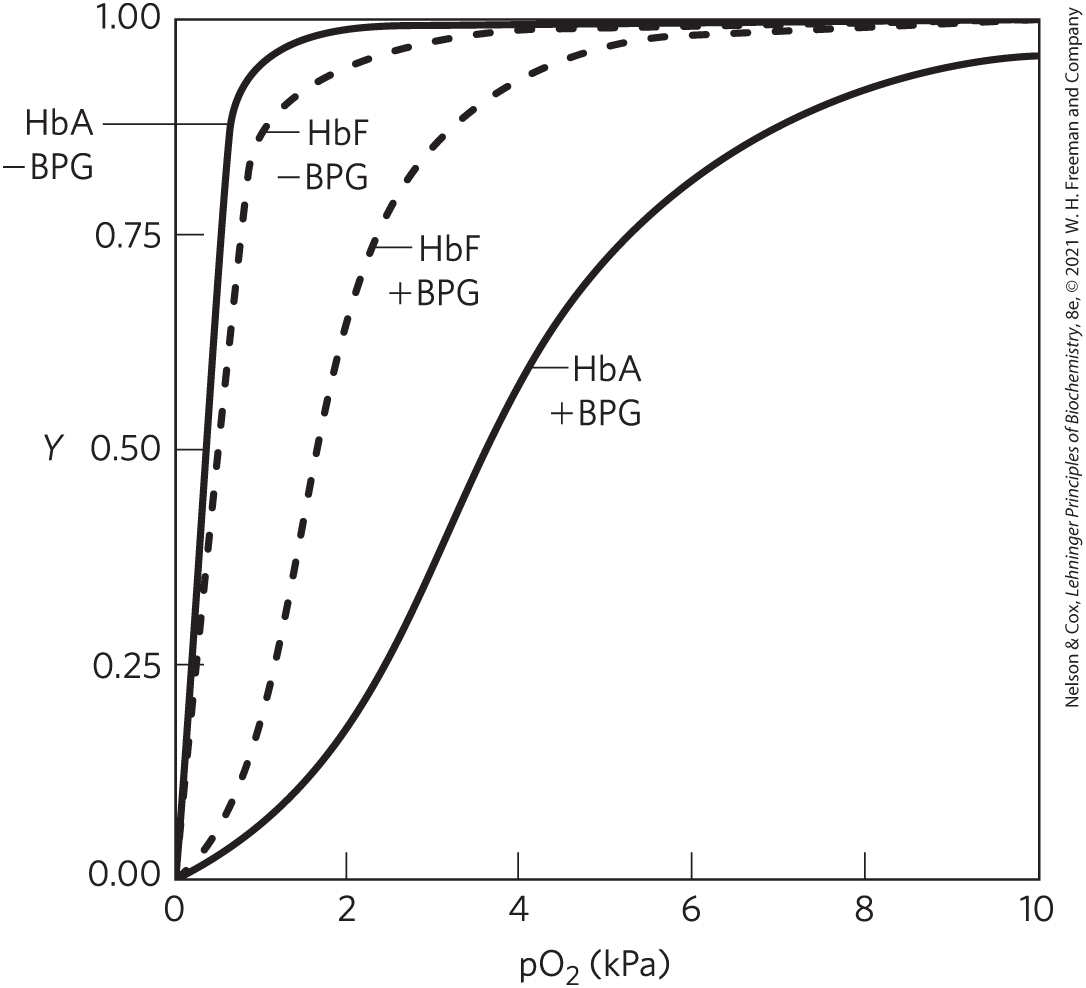
- What is the effect of BPG on the affinity of hemoglobin? How can this information be used to explain the different affinities of fetal and maternal hemoglobin?
10. Hemoglobin Variants There are almost 500 naturally occurring variants of hemoglobin. Most are the result of a single amino acid substitution in a globin polypeptide chain. Some variants produce clinical illness, though not all variants have deleterious effects. A brief sample of hemoglobin variants is shown here.
- HbS (sickle cell Hb): substitutes a Val for a Glu on the surface
- Hb Cowtown: eliminates an ion pair involved in T-state stabilization
- Hb Memphis: substitutes one uncharged polar residue for another of similar size on the surface
- Hb Bibba: substitutes a Pro for a Leu involved in an α helix
- Hb Milwaukee: substitutes a Glu for a Val
- Hb Providence: substitutes an Asn for a Lys that normally projects into the central cavity of the tetramer
- Hb Philly: substitutes a Phe for a Tyr, disrupting hydrogen bonding at the interface
Select the hemoglobin variants that are described by each statement.
- The Hb variant least likely to cause pathological symptoms
- The variant(s) most likely to show pI values different from that of HbA on an isoelectric focusing gel
- The variant(s) most likely to show a decrease in BPG binding and an increase in the overall affinity of the hemoglobin for oxygen
11. Oxygen Binding and Hemoglobin Structure A team of biochemists uses genetic engineering to modify the interface region between hemoglobin subunits. The resulting hemoglobin variants exist in solution primarily as αβ dimers (few, if any, tetramers form). Are these variants likely to bind oxygen more weakly or more tightly? Explain your answer.
12. Reversible (and Tight) Binding to an Antibody An antibody with high affinity for its antigen has a in the low nanomolar range. Assume an antibody binds an antigen with a of . Calculate the antigen concentration when Y, the fraction of binding sites occupied by the ligand, is
- 0.4,
- 0.5,
- 0.8,
- 0.9.
13. Using Antibodies to Probe Structure-Function Relationships in Proteins A monoclonal antibody binds to G-actin but not to F-actin. What does this tell you about the epitope recognized by the antibody?
14. The Immune System and Vaccines Some pathogens have developed mechanisms to evade the immune system, making it difficult or impossible to develop effective vaccines against them.
- African sleeping sickness is caused by a protozoan called Trypanosoma brucei, carried by the tsetse fly. The trypanosome surface is dominated by one coat protein, the variable surface glycoprotein (VSG). The trypanosome genome encodes over 1,000 different versions of VSG. All of the cells in an initial infection feature the same VSG coat on their surfaces, and this is readily recognized as foreign by the immune system. However, an individual trypanosome in the broader population will switch and randomly begin expressing a different variant of the VSG coat. All the descendants of that cell will have the new and different protein on their surface. As the population with the second VSG coat increases, an individual cell will then switch to a third VSG protein coat, and so on.
- The human immunodeficiency virus (HIV) has an error-prone system for replicating its genome, effectively introducing mutations at an unusually high rate. Many of the mutations affect the viral protein coat. Describe how each pathogen can survive the immune response of its host.
15. How We Become a “Stiff” When a vertebrate dies, its muscles stiffen as they are deprived of ATP, a state called rigor mortis. Using your knowledge of the catalytic cycle of myosin in muscle contraction, explain the molecular basis of the rigor state.
16. Sarcomeres from Another Point of View The symmetry of thick and thin filaments in a sarcomere is such that six thin filaments ordinarily surround each thick filament in a hexagonal array. Match each cross section (transverse cut) image of a sarcomere with the correct viewpoint.
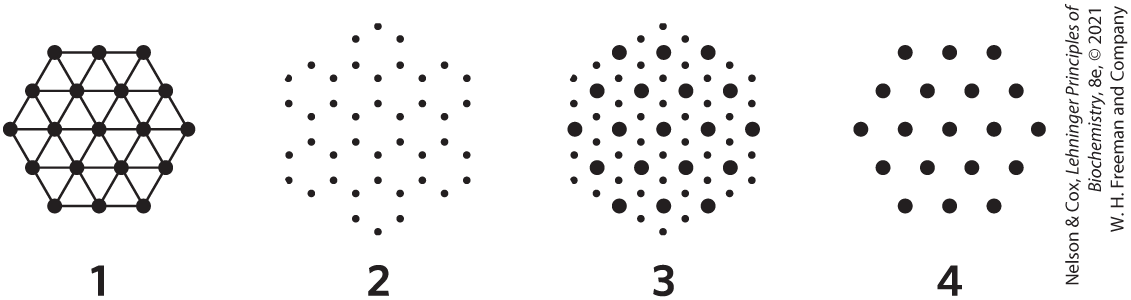
- at the M line
- through the I band
- through the dense region of the A band
- through the less dense region of the A band, adjacent to the M line (see Fig. 5-28b, c)
 Reversible Ligand Binding IV Exposure to carbon monoxide can lead to unconsciousness and ultimately death. Suffocation occurs when hemoglobin is half-saturated with CO — that is, when only two of every four oxygen-binding sites are occupied with CO. Explain why death may occur at this point, even though half of the oxygen-binding sites are still available to transport . (Hint: See
Reversible Ligand Binding IV Exposure to carbon monoxide can lead to unconsciousness and ultimately death. Suffocation occurs when hemoglobin is half-saturated with CO — that is, when only two of every four oxygen-binding sites are occupied with CO. Explain why death may occur at this point, even though half of the oxygen-binding sites are still available to transport . (Hint: See BIOCHEMISTRY ONLINE
17. IgG and Lysozyme Crystal Structure To fully appreciate how proteins function in a cell, it is helpful to have a three-dimensional view of how proteins interact with other cellular components. Fortunately, this is possible using online protein databases and three-dimensional molecular viewing utilities such as JSmol, a free and user-friendly molecular viewer that is compatible with most browsers and operating systems.
In this exercise, examine the interactions between the enzyme lysozyme and the Fab portion of the antilysozyme antibody. Use the PDB identifier 1FDL to explore the structure of the IgG1 Fab fragment–lysozyme complex (antibody-antigen complex). To answer the questions, use the information on the Structure Summary page at the Protein Data Bank (www.rcsb.org), and view the structure using JSmol or a similar viewer.
- Which chains in the three-dimensional model correspond to the antibody fragment, and which correspond to the antigen, lysozyme?
- What type of secondary structure predominates in this Fab fragment?
- How many amino acid residues are in the heavy and light chains of the Fab fragment? In lysozyme? Estimate the percentage of the lysozyme that interacts with the antigen-binding site of the antibody fragment.
- Identify the specific amino acid residues in lysozyme and in the variable regions of the Fab heavy and light chains that are situated at the antigen-antibody interface. Are the residues contiguous in the primary sequence of the polypeptide chains?
18. Exploring Antibodies in the Protein Data Bank The PDB-101 Molecule of the Month article on “Antibodies” (http://pdb101.rcsb.org/motm/21) summarizes what you have read in this chapter regarding antibody structure and function. To paraphrase the article, a variety of antibodies, on the order of one hundred million different types, are always circulating in our bloodstream, searching for foreign invaders to attack. Once an invader is discovered, the antibody binds the invader with its flexible arms, containing the Fab region. Thin, flexible chains connect these flexible arms to the antibody base, called the Fc region. This base determines which class the antibody belongs to, as some antibodies have four or ten binding sites due to their structural formation.
- How many specific antigen-binding sites are there on the first immunoglobulin image (derived from PDB ID 1IGT) in the article?
- When a virus enters your lungs, how long does it take for you to produce one or more antibodies that bind to it?
- Approximately how many types of different antibodies are present in your blood?
- Explore the structure of the immunoglobulin molecule (PDB ID 1IGT) by clicking the link in the article or by using a search engine to find the structure summary for PDB ID 1IGT. Use one of the 3D viewers on the PDB site to view a ribbon structure for this immunoglobulin. Identify the two light chains and two heavy chains (use the viewer controls to distinguish them by color).
DATA ANALYSIS PROBLEM
19. Protein Function During the 1980s, the structures of actin and myosin were known only at the resolution shown in Figure 5-26a. Although researchers knew that the globular head portion of myosin bound to actin and hydrolyzed ATP, there was a substantial debate about where in the myosin molecule the contractile force was generated. At the time, two competing models were proposed for the mechanism of force generation in myosin.
In the “hinge” model, the head bound to actin, but the pulling force was generated by contraction of the “hinge region” in the myosin tail. The hinge region is in the heavy meromyosin portion of the myosin molecule; this is roughly the point labeled “Two supercoiled α helices” in Figure 5-26. In the “S1” model (S1 being a name used to describe the head), the pulling force was generated in the S1 “head” itself and the tail was just for structural support.
Many experiments were performed but provided no conclusive evidence. Then, in 1987, James Spudich and his colleagues at Stanford University published a study that, although not conclusive, went a long way toward resolving this controversy.
Recombinant DNA techniques were not sufficiently developed to address this issue in vivo, so Spudich and colleagues used an interesting in vitro motility assay. The alga Nitella has extremely long cells, often several centimeters long and about 1 mm in diameter. These cells have actin fibers that run along their long axes, and the cells can be cut open along their length to expose the actin fibers. Spudich and his group had observed that plastic beads coated with myosin would “walk” along these fibers in the presence of ATP, just as myosin would do in contracting muscle.
For these experiments, the researchers used a more well-defined method for attaching the myosin to the beads. The “beads” were clumps of killed bacterial (Staphylococcus aureus) cells. These cells have a protein on their surface that binds to the Fc region of antibody molecules (Fig. 5-20a). The antibodies, in turn, bind to several (unknown) places along the tail of the myosin molecule. When bead-antibody-myosin complexes were prepared with intact myosin molecules, they would move along Nitella actin fibers in the presence of ATP.
- Sketch a diagram showing what a bead-antibody-myosin complex might look like at the molecular level.
- Why was ATP required for the beads to move along the actin fibers?
- Spudich and coworkers used antibodies that bound to the myosin tail. Why would this experiment have failed if they had used an antibody that bound to the part of S1 that normally bound to actin? Why would this experiment have failed if they had used an antibody that bound to actin?
To help focus on the part of myosin responsible for force production, Spudich and colleagues used trypsin to produce two partial myosin molecules: heavy meromyosin (HMM) and light meromyosin (LMM), by cleavage of a single specific peptide bond in the myosin tail. Additional incubation with trypsin produced an additional cleavage, eliminating more of the tail and the hinge region to generate short heavy meromyosin (SHMM).
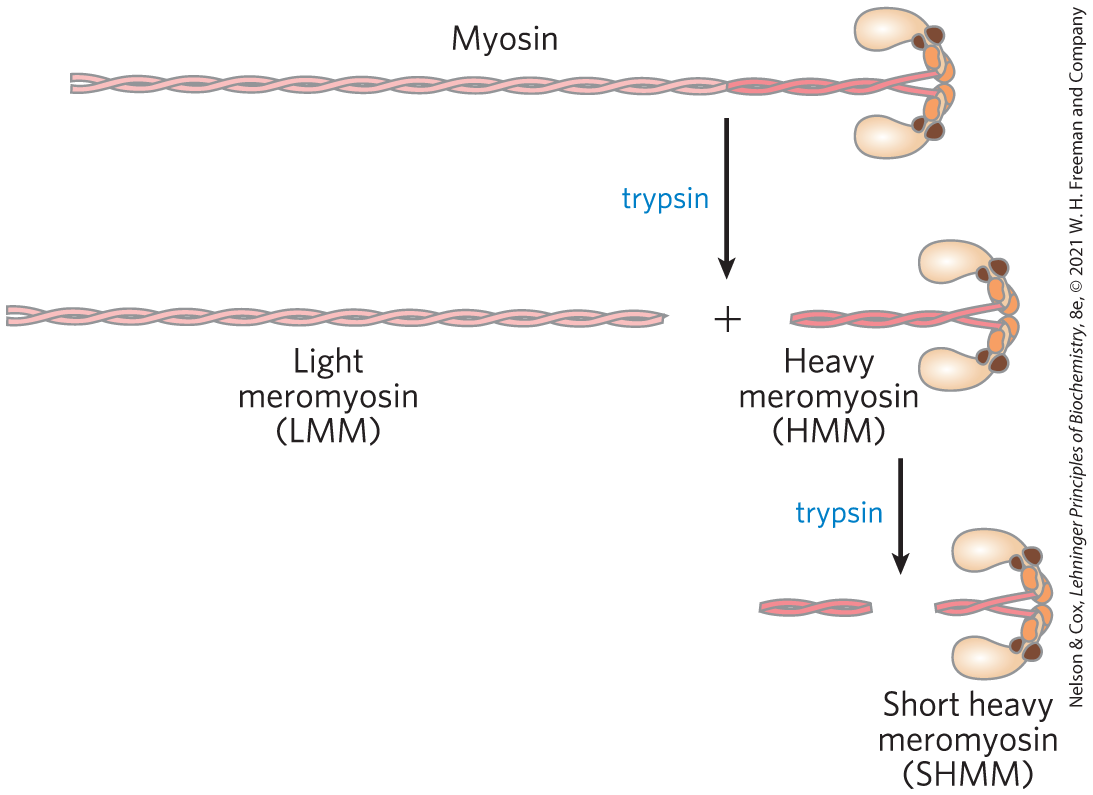
- Why might trypsin attack this peptide bond first, rather than other peptide bonds in myosin?
Spudich and colleagues prepared bead-antibody-myosin complexes with varying amounts of myosin, HMM, or SHMM and measured their speed of movement along Nitella actin fibers in the presence of ATP. The graph below sketches their results.
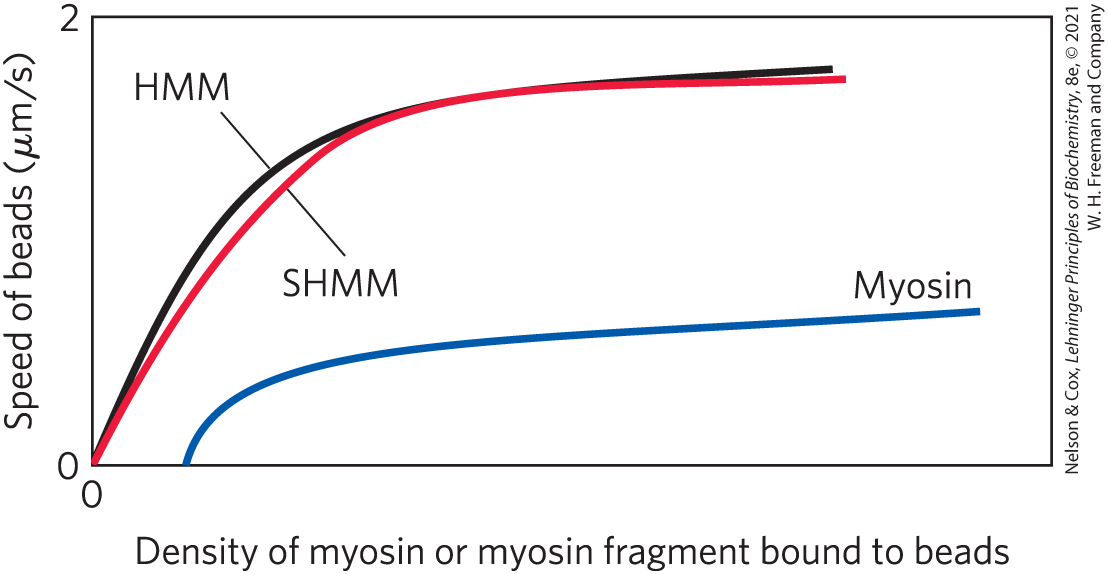
- Which model (“S1” or “hinge”) is consistent with these results? Explain your reasoning.
- Provide a plausible explanation for the increased speed of the beads with increasing myosin density.
- Provide a plausible explanation for the plateauing of the speed of the beads at high myosin density.
Reference
- Hynes, T.R., S.M. Block, B.T. White, and J.A. Spudich. 1987. Movement of myosin fragments in vitro: domains involved in force production. Cell 48:953–963.