How is a concentration gradient of protons transformed into ATP? We have seen that electron transfer releases, and the proton-motive force conserves, more than enough free energy (about 190 kJ) per “mole” of electron pairs to drive the formation of a mole of ATP, which requires about 50 kJ (p. 480). Mitochondrial oxidative phosphorylation therefore poses no thermodynamic problem. But what is the chemical mechanism that couples proton flux with phosphorylation?
In the Chemiosmotic Model, Oxidation and Phosphorylation Are Obligately Coupled
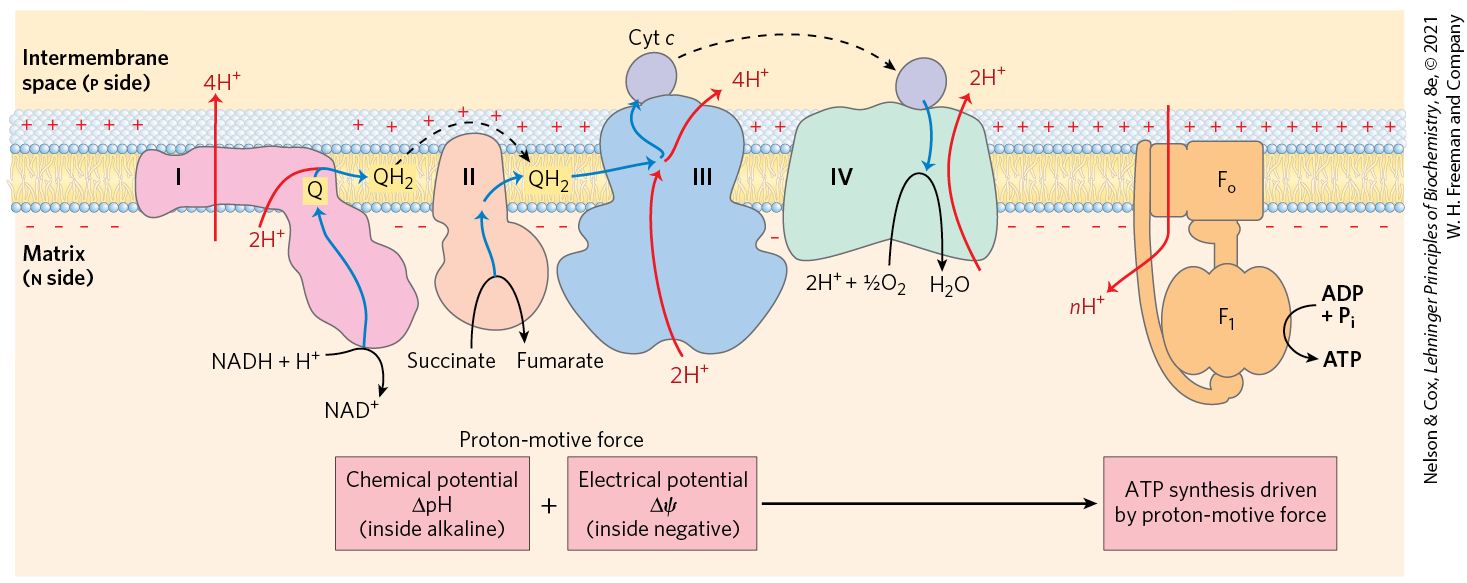
The chemiosmotic model, proposed by Peter Mitchell, is the paradigm for energy coupling. According to the model (Fig. 19-19), the electrochemical energy inherent in the difference in proton concentration and the separation of charge across the inner mitochondrial membrane — the proton-motive force — drives the synthesis of ATP as protons flow passively back into the matrix through a proton pore in ATP synthase. To emphasize this crucial role of the proton-motive force, the equation for ATP synthesis is sometimes written
(19-10)
FIGURE 19-19 Chemiosmotic model. In this simple representation of the chemiosmotic theory applied to mitochondria, electrons from NADH and other oxidizable substrates pass through a chain of carriers arranged asymmetrically in the inner membrane. Electron flow is accompanied by proton transfer across the membrane, producing both a chemical gradient and an electrical gradient , which, combined, create the proton-motive force. The inner mitochondrial membrane is impermeable to protons; protons can reenter the matrix only through proton-specific channels . The proton-motive force that drives protons back into the matrix provides the energy for ATP synthesis, catalyzed by the complex associated with .
Peter Mitchell, 1920–1992
Mitchell used the term “chemiosmotic” to describe enzymatic reactions that involve, simultaneously, a chemical reaction and a transport process, and the overall process is sometimes referred to as “chemiosmotic coupling.” Here, “coupling” refers to the obligate connection between mitochondrial ATP synthesis and electron flow through the respiratory chain; neither of the two processes can proceed without the other. The operational definition of coupling is shown in Figure 19-20. When isolated mitochondria are suspended in a buffer containing ADP, , and an oxidizable substrate such as succinate, three easily measured processes occur: (1) the substrate is oxidized (succinate yields fumarate), (2) is consumed, and (3) ATP is synthesized. Oxygen consumption and ATP synthesis depend on the presence of an oxidizable substrate (succinate in this case) as well as ADP and .
FIGURE 19-20 Coupling of electron transfer and ATP synthesis in mitochondria. In experiments to demonstrate coupling, mitochondria are suspended in a buffered medium, and an electrode monitors consumption. At intervals, samples are removed and assayed for the presence of ATP. (a) Addition of ADP and alone results in little or no increase in either respiration ( consumption; black) or ATP synthesis (red). When succinate is added, respiration begins immediately, and ATP is synthesized. Addition of cyanide , which blocks electron transfer between cytochrome oxidase (Complex IV) and , inhibits both respiration and ATP synthesis. (b) Mitochondria provided with succinate respire and synthesize ATP only when ADP and are added. Subsequent addition of venturicidin or oligomycin, inhibitors of ATP synthase, blocks both ATP synthesis and respiration. Dinitrophenol (DNP) is an uncoupler, allowing respiration to continue without ATP synthesis.
Because substrate oxidation drives ATP synthesis, inhibitors of electron transfer block ATP synthesis (Fig. 19-20a). The converse is also true: inhibition of ATP synthesis blocks electron transfer in intact mitochondria. When isolated mitochondria are given and oxidizable substrates, but not ADP (Fig. 19-20b), no ATP synthesis can occur and electron transfer to does not proceed. Henry Lardy, who pioneered the use of antibiotics to explore mitochondrial function, showed coupling of oxidation and phosphorylation by using oligomycin and venturicidin. These toxic antibiotics bind to the ATP synthase in mitochondria, inhibiting both ATP synthesis and the transfer of electrons through the chain of carriers to (Fig. 19-20b). As oligomycin does not interact with the electron carriers, it follows that electron transfer and ATP synthesis are obligately coupled: neither reaction occurs without the other.
Henry Lardy, 1917–2010
Chemiosmotic theory readily explains the dependence of electron transfer on ATP synthesis in mitochondria. When the flow of protons into the matrix through the proton channel of ATP synthase is blocked (with oligomycin, for example), no path exists for the return of protons to the matrix, and the continued extrusion of protons driven by the activity of the respiratory chain generates a large proton gradient. The proton-motive force builds up until the cost (free energy) of pumping protons out of the matrix against this gradient equals or exceeds the energy released by the transfer of electrons from NADH to . At this point electron flow must stop; the free energy for the overall process of electron flow coupled to proton pumping becomes zero, and the system is at equilibrium.
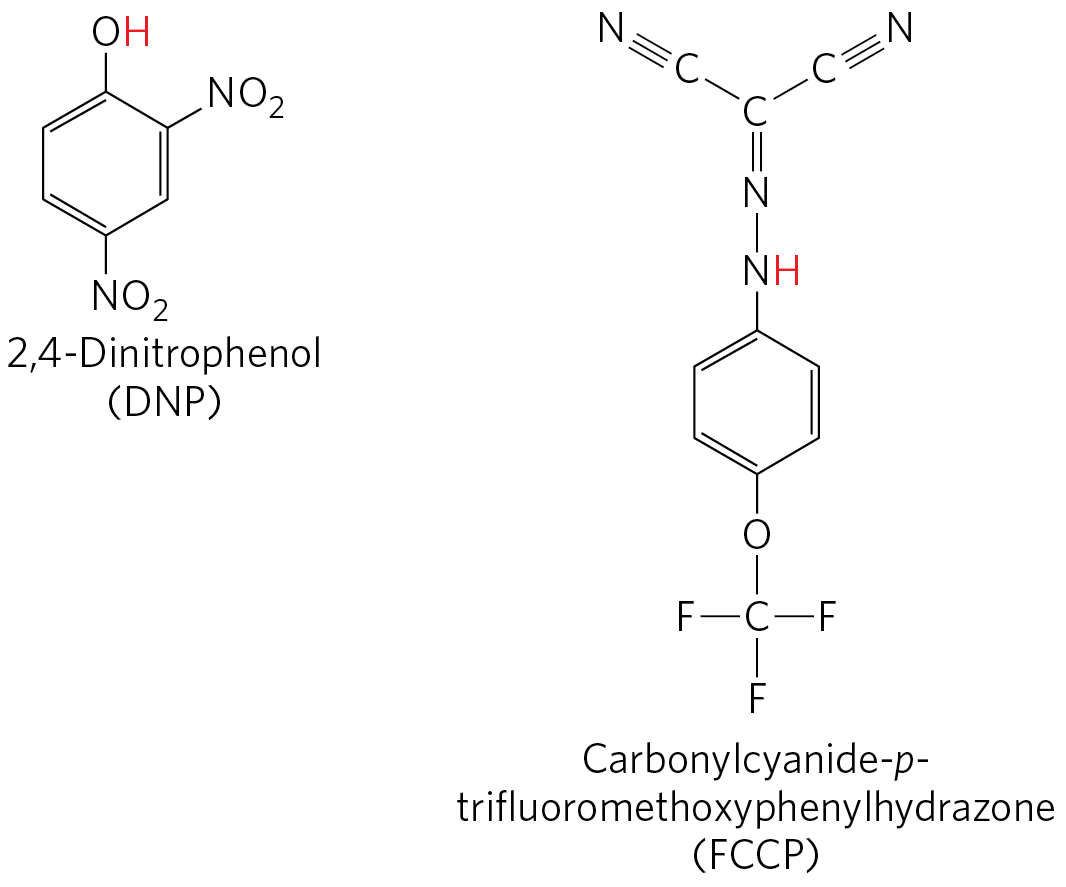
Certain conditions and reagents, however, can uncouple oxidation from phosphorylation. When intact mitochondria are disrupted by treatment with detergent or by physical shear, the resulting membrane fragments can still catalyze electron transfer from succinate or NADH to , but no ATP synthesis is coupled to this respiration. Certain chemical compounds cause uncoupling without physically disrupting mitochondrial structure. Chemical uncouplers include 2,4-dinitrophenol (DNP) and carbonylcyanide-p-trifluoromethoxyphenylhydrazone (FCCP) (Table 19-4; Fig. 19-21), weak acids with hydrophobic properties that permit them to diffuse readily across mitochondrial membranes. After entering the matrix in the protonated form, they can release a proton, thus dissipating the proton gradient. Resonance stabilization delocalizes the charge on the anionic forms, making them sufficiently hydrophobic to diffuse back across the membrane, where they can pick up a proton and repeat the process. Ionophores such as valinomycin (see Fig. 11-43) allow inorganic ions to pass easily through membranes. Ionophores uncouple electron transfer from oxidative phosphorylation by dissipating the electrical contribution to the electrochemical gradient across the mitochondrial membrane.
FIGURE 19-21 Two chemical uncouplers of oxidative phosphorylation. Both DNP and FCCP have a dissociable proton (red) and are very hydrophobic. They carry protons across the inner mitochondrial membrane, dissipating the proton gradient. Both also uncouple photophosphorylation.
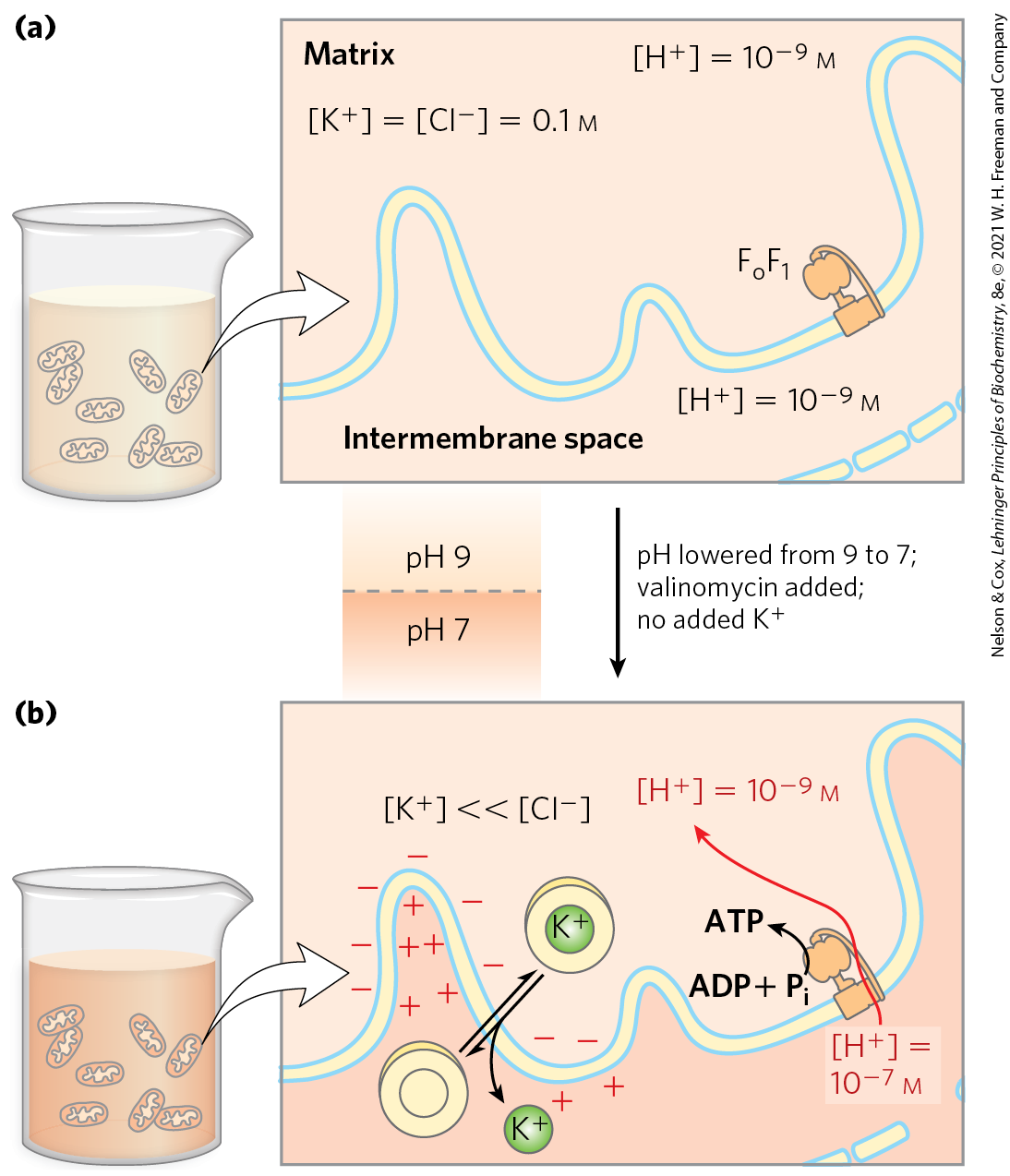
A prediction of the chemiosmotic theory is that, because the role of electron transfer in mitochondrial ATP synthesis is simply to pump protons to create the electrochemical potential of the proton-motive force, an artificially created proton gradient should be able to replace electron transfer in driving ATP synthesis. This has been experimentally confirmed (Fig. 19-22). In the absence of an oxidizable substrate, the proton-motive force alone suffices to drive ATP synthesis.
FIGURE 19-22 Evidence for the role of a proton gradient in ATP synthesis. An artificially imposed electrochemical gradient can drive ATP synthesis in the absence of an oxidizable substrate as electron donor. In this two-step experiment, (a) isolated mitochondria are first incubated in a pH 9 buffer containing 0.1 m KCl. Slow leakage of buffer and KCl into the mitochondria eventually brings the matrix into equilibrium with the surrounding medium. No oxidizable substrates are present. (b) Mitochondria are now removed from the pH 9 buffer and resuspended in pH 7 buffer containing valinomycin but no KCl. The change in buffer creates a difference of two pH units across the inner mitochondrial membrane. The outward flow of , carried by valinomycin down the ion concentration gradient without a counterion, creates a charge imbalance across the membrane (matrix negative). The sum of the chemical potential provided by the pH difference and the electrical potential provided by the separation of charges is a proton-motive force large enough to support ATP synthesis in the absence of an oxidizable substrate.
ATP Synthase Has Two Functional Domains, and
Mitochondrial ATP synthase is an F-type ATPase (see Fig. 11-40b) similar in structure and mechanism to the ATP synthases of bacteria and (as we will see in Chapter 20) chloroplasts. This large enzyme complex of the inner mitochondrial membrane catalyzes the formation of ATP from ADP and , driven by the flow of protons from the p to the n side of the membrane (Eqn 19-10). ATP synthase, also called Complex V to relate it to the electron-transfer complexes described in the last section, has two distinct components. These are , a peripheral membrane protein, and (o denoting oligomycin-sensitive), which is integral to the membrane. , the first factor recognized as essential for oxidative phosphorylation, was identified and purified by Efraim Racker and his colleagues in the early 1960s.
In the laboratory, small membrane vesicles formed from inner mitochondrial membranes carry out ATP synthesis coupled to electron transfer. When is gently extracted, the “stripped” vesicles still contain intact respiratory chains and the portion of ATP synthase. The vesicles can catalyze electron transfer from NADH to but cannot produce a proton gradient: has a proton pore through which protons leak as fast as they are pumped by electron transfer, and without a proton gradient the -depleted vesicles cannot make ATP. Isolated catalyzes ATP hydrolysis (the reversal of synthesis) and was therefore originally called ATPase. When purified is added back to the depleted vesicles, it reassociates with , plugging its proton pore and restoring the membrane’s capacity to couple electron transfer and ATP synthesis.
ATP Is Stabilized Relative to ADP on the Surface of
Isotope exchange experiments using purified reveal an extraordinary fact about the enzyme’s catalytic mechanism: on the enzyme surface, the reaction is readily reversible — the free-energy change for ATP synthesis is close to zero. When ATP is hydrolyzed by in the presence of -labeled water, the released contains an atom. Careful measurement of the content of formed in vitro by -catalyzed hydrolysis of ATP reveals that the has not one but three or four atoms (Fig. 19-23). This indicates that the terminal pyrophosphate bond in ATP is cleaved and re-formed repeatedly before leaves the enzyme surface. This exchange reaction occurs in unenergized complexes (with no proton gradient) and with isolated — the exchange does not require the input of energy.
FIGURE 19-23 Catalytic mechanism of . (a) An -exchange experiment. solubilized from mitochondrial membranes is incubated with ATP in the presence of -labeled water. At intervals, a sample of the solution is withdrawn and analyzed for the incorporation of into the produced from ATP hydrolysis. In minutes, the contains three or four atoms, indicating that both ATP hydrolysis and ATP synthesis have occurred several times during the incubation. (b) The likely transition state complex for ATP hydrolysis and synthesis by ATP synthase. The α subunit is shown in gray, β in purple. The positively charged residues and coordinate two oxygens of the pentavalent phosphate intermediate; interacts with a third oxygen, and the ion further stabilizes the intermediate. The blue sphere represents the leaving group . These interactions result in ready equilibration of ATP and in the active site. [(b) Data from PDB ID 1BMF, J. P. Abrahams et al., Nature 370:621, 1994.]
Kinetic studies of the initial rates of ATP synthesis and hydrolysis confirm the conclusion that for ATP synthesis on the enzyme is near zero. From the measured rates of hydrolysis and synthesis , the calculated equilibrium constant for the reaction
is
From this , the calculated apparent is close to zero. This is much different from the of about for the hydrolysis of ATP free in solution (i.e., not on the enzyme surface).
What accounts for the huge difference? ATP synthase stabilizes ATP relative to by binding ATP more tightly, releasing enough energy to counterbalance the cost of making ATP. Careful measurements of the binding constants show that binds ATP with very high affinity and ADP with much lower affinity . The difference in corresponds to a difference of about 40 kJ/mol in binding energy, and this binding energy drives the equilibrium toward formation of the product ATP.
The Proton Gradient Drives the Release of ATP from the Enzyme Surface
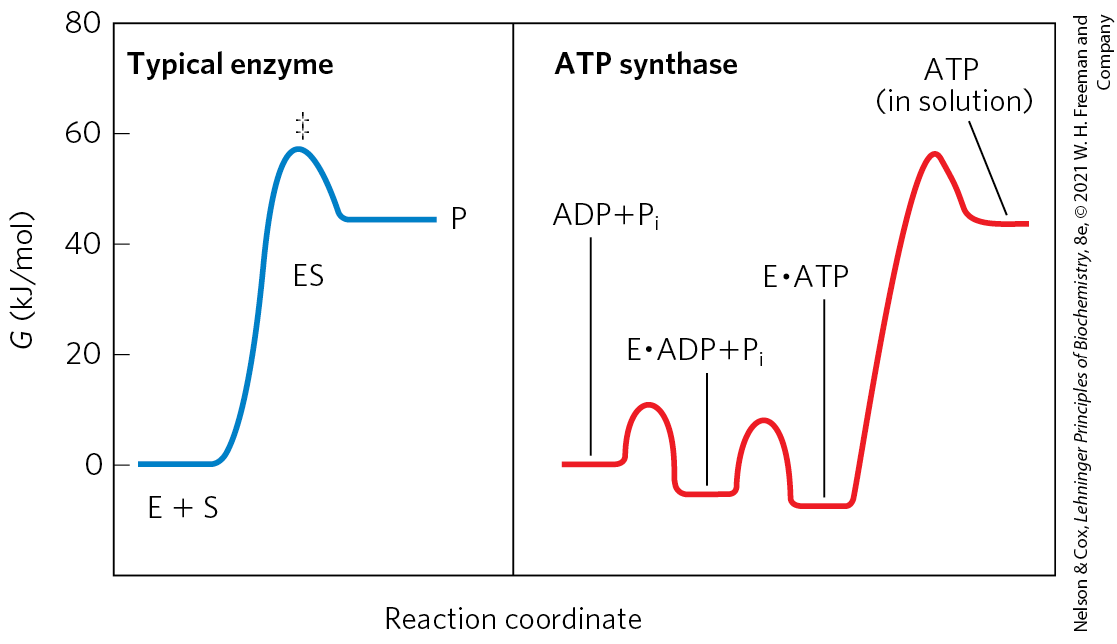
Although ATP synthase equilibrates ATP with , in the absence of a proton gradient the newly synthesized ATP does not leave the surface of the enzyme. Effectively, the enzyme cannot turn over and synthesize a second molecule of ATP. It is the proton gradient that causes the enzyme to release the ATP formed on its surface. The reaction coordinate diagram of the process (Fig. 19-24) illustrates the difference between the mechanism of ATP synthase and that of many other enzymes that catalyze endergonic reactions.
FIGURE 19-24 Reaction coordinate diagrams for ATP synthase and for a more typical enzyme. In a typical enzyme-catalyzed reaction (left), reaching the transition state between substrate and product is the major energy barrier to overcome. In the reaction catalyzed by ATP synthase (right), release of ATP from the enzyme, not formation of ATP, is the major energy barrier. The free-energy change for the formation of ATP from ADP and in aqueous solution is large and positive, but on the enzyme surface, the very tight binding of ATP provides sufficient binding energy to bring the free energy of the enzyme-bound ATP close to that of , so the reaction is readily reversible. The equilibrium constant is near 1. The free energy required for the release of ATP is provided by the proton-motive force.
For the continued synthesis of ATP, the enzyme must cycle between a form that binds ATP very tightly and a form that releases ATP. Chemical and crystallographic studies of the ATP synthase have revealed the structural basis for this alternation in function.
Each β Subunit of ATP Synthase Can Assume Three Different Conformations
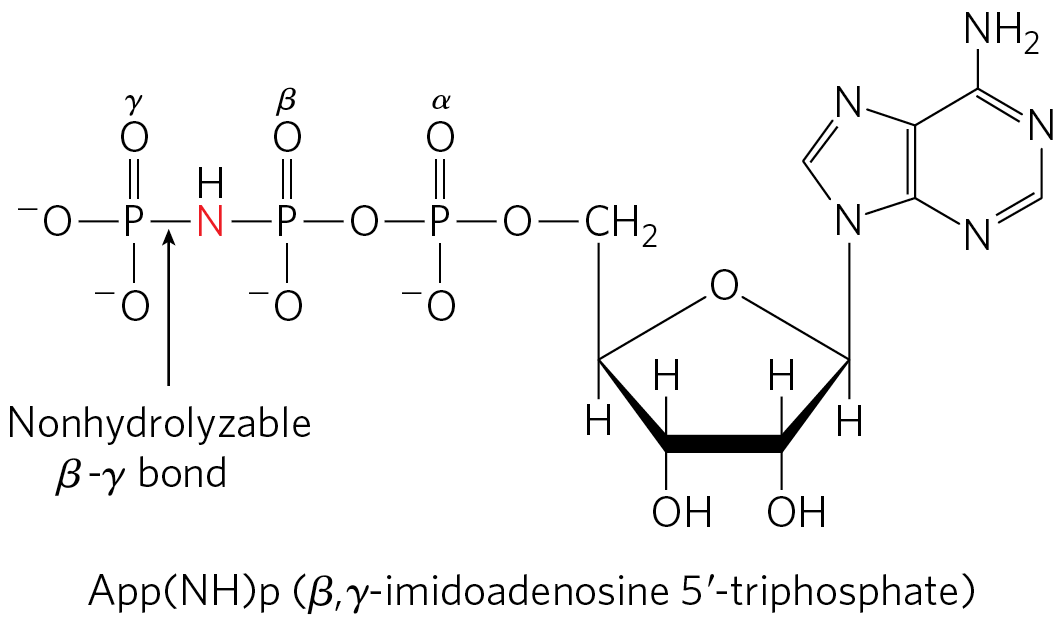
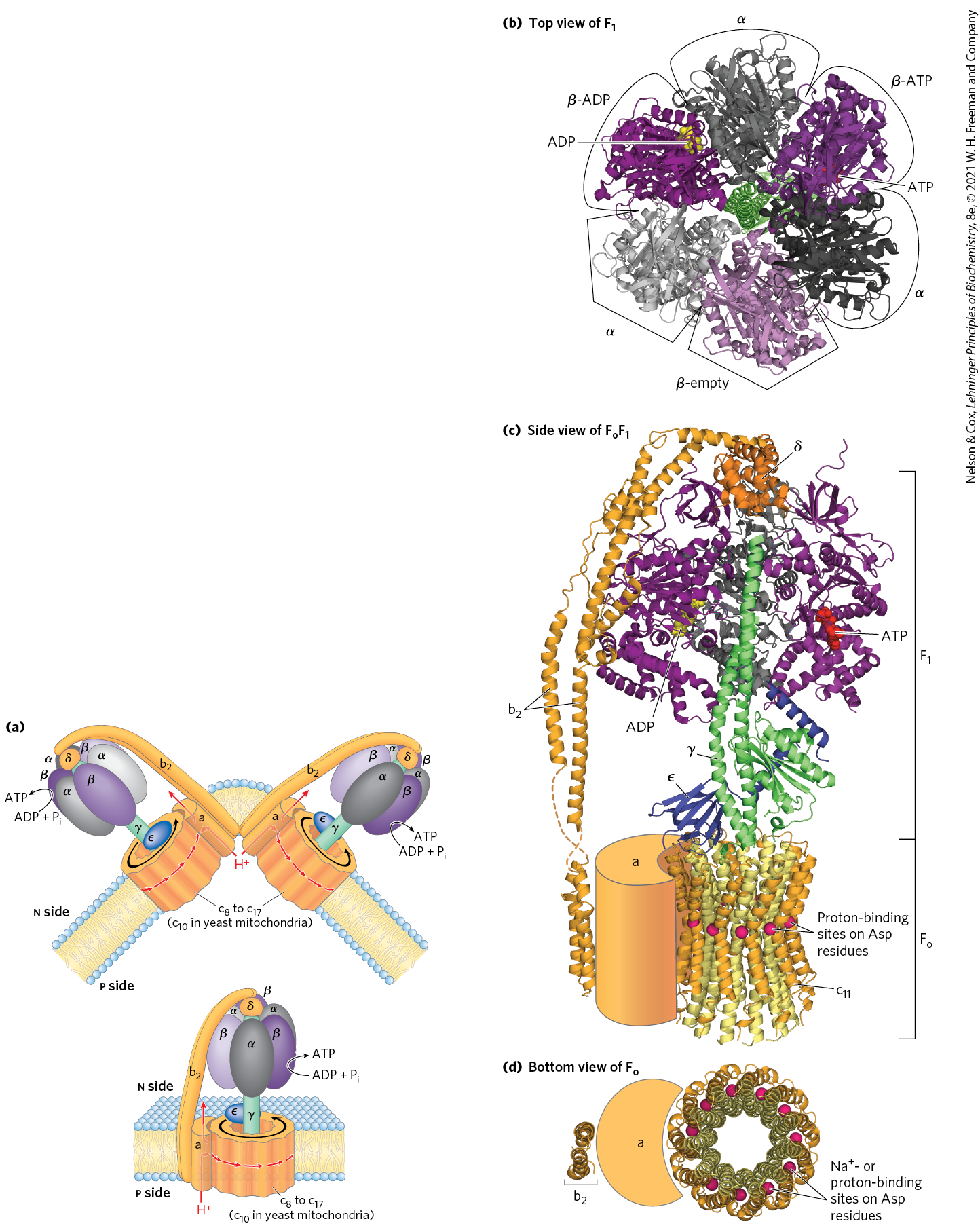
Mitochondrial has nine subunits of five different types, with the composition . Each of the three β subunits has one catalytic site for ATP synthesis. The crystallographic determination of the structure by John E. Walker and colleagues revealed structural details that help explain the catalytic mechanism of the enzyme. The knoblike portion of is a flattened sphere, 8 nm by 10 nm, consisting of alternating α and β subunits arranged like the sections of an orange (Fig. 19-25a–d). Although the amino acid sequences of the three β subunits are identical, their conformations differ. The conformational differences extend to differences in their ATP/ADP-binding sites. When the protein is crystallized in the presence of ADP and App(NH)p, a close structural analog of ATP that cannot be hydrolyzed by the ATPase activity of , the binding site of one of the three β subunits is filled with App(NH)p, the second is filled with ADP, and the third is empty. The corresponding β subunit conformations are designated β-ATP, β-ADP, and β-empty (Fig. 19-25b). This difference in nucleotide binding among the three subunits is critical to the mechanism of the complex. The polypeptides that make up the stalk in the crystal structure are asymmetrically arranged. One domain of the single γ subunit makes up a central shaft that passes through . Another globular domain of γ helps to stabilize the β-empty conformation in a β subunit it is transiently associated with (Fig. 19-25c).
FIGURE 19-25 Mitochondrial ATP synthase complex. (a) A cartoon representation of the complex. The dimeric form is found in eukaryotic mitochondria. The monomeric form is observed in bacteria. (b) viewed from above (that is, from the n side of the membrane), showing the three β (shades of purple) and three α (shades of gray) subunits and the central shaft (γ subunit, green). Each β subunit, near its interface with the neighboring α subunit, has a nucleotide-binding site critical to the catalytic activity. The single γ subunit associates primarily with one of the three pairs, forcing each of the three β subunits into slightly different conformations, with different nucleotide-binding sites. In the crystalline enzyme, one subunit, β-ADP, has ADP (yellow) in its binding site; the next, β-ATP, has ATP (red); and the third, β-empty, has no bound nucleotide. (c) The entire enzyme viewed from the side (in the plane of the membrane). The portion has three α subunits and three β subunits arranged like the segments of an orange around a central shaft, the γ subunit (green). (Two α subunits and one β subunit have been omitted to reveal the γ subunit and the binding sites for ATP and ADP on the β subunits.) The δ subunit confers oligomycin sensitivity on the ATP synthase, and the ε subunit may serve to inhibit the enzyme’s ATPase activity under some circumstances. The subunit consists of one a subunit and two b subunits, which anchor the complex in the membrane and act as a stator (the stationary part of a rotary system), holding the α and β subunits in place. also includes the c ring, made up of a number (8 to 17, depending on the species) of identical c subunits, small, hydrophobic proteins. The c ring and the a subunit interact to provide a transmembrane path for protons. Each of the c subunits in has a critical Asp residue near the middle of the membrane, which undergoes protonation/deprotonation during the catalytic cycle of the ATP synthase. Shown here is the homologous ring of the of Ilyobacter tartaricus, for which the structure is well established. The -binding sites, which correspond to the proton-binding sites of the complex, are shown with their bound ions (red spheres). (d) A view of perpendicular to the membrane. As in (c), red spheres represent the or proton-binding sites in Asp residues. [(a) Information from W. Kühlbrandt and K. M. Davies, Trends Biochem. Sci. 41:106, 2016. (b, c, d) Data from : PDB ID 1BMF, J. P. Abrahams et al., Nature 370:621, 1994; PDB ID 1JNV, A. C. Hausrath et al., J. Biol. Chem. 276:47,227, 2001; PDB ID 2A7U, S. Wilkens et al., Biochemistry 44:11,786, 2005; PDB ID 2CLY, V. Kane Dickson et al., EMBO J. 25:2911, 2006; : PDB ID 1B9U, O. Dmitriev et al., J. Biol. Chem. 274:15,598, 1999; c ring: PDB ID 1YCE, T. Meier et al., Science 308:659, 2005.]
The complex, with its proton pore, is composed of three subunits, a, b, and c, in the proportion , where n ranges from 8 to 17, depending on the species. Subunit c is a small , very hydrophobic polypeptide, consisting almost entirely of two transmembrane helices, with a small loop extending from the matrix side of the membrane. The crystal structure of the yeast shows 10 c subunits, each with two transmembrane helices roughly perpendicular to the plane of the membrane and arranged in two concentric circles to create the c ring. The inner circle is made up of the amino-terminal helices of each c subunit; the outer circle, about 55 Å in diameter, is made up of the carboxyl-terminal helices. The c subunits in the c ring rotate together as a unit around an axis perpendicular to the membrane. The and γ subunits of form a leg-and-foot that projects from the bottom (membrane) side of and stands firmly on the ring of c subunits. The a subunit consists of several hydrophobic helices that span the membrane in close association with one of the c subunits in the c ring.
Rotational Catalysis Is Key to the Binding-Change Mechanism for ATP Synthesis
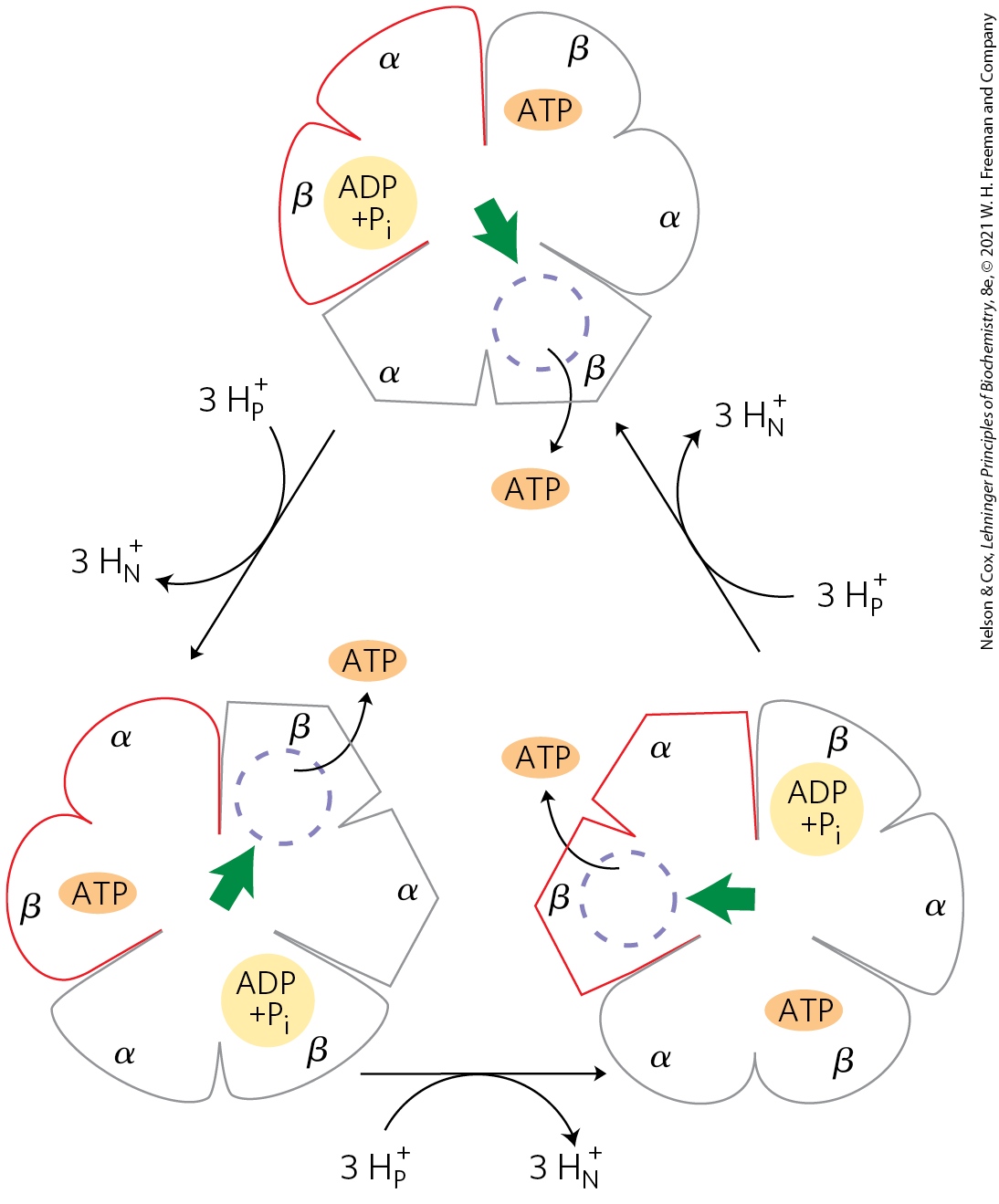
On the basis of detailed kinetic and binding studies of the reactions catalyzed by , Paul Boyer proposed a rotational catalysis mechanism in which the three active sites of take turns catalyzing ATP synthesis (Fig. 19-26). A given β subunit starts in the β-ADP conformation, which binds ADP and from the surrounding medium. The subunit now changes conformation, assuming the β-ATP form that tightly binds and stabilizes ATP, bringing about the ready equilibration of with ATP on the enzyme surface. Finally, the subunit changes to the β-empty conformation, which has very low affinity for ATP, and the newly synthesized ATP leaves the enzyme surface. Another round of catalysis begins when this subunit again assumes the β-ADP form and binds ADP and .
FIGURE 19-26 Binding-change model for ATP synthase. The complex has three nonequivalent adenine nucleotide–binding sites, one for each pair of α and β subunits. At any given moment, one of these sites is in the β-ATP conformation (which binds ATP tightly), a second is in the β-ADP (loose-binding) conformation, and a third is in the β-empty (very-loose-binding) conformation. In this view from the n side, the proton-motive force causes rotation of the central shaft — the γ subunit, shown as a green arrowhead — which comes into contact with each subunit pair in succession. This produces a cooperative conformational change in which the β-ATP site is converted to the β-empty conformation, and ATP dissociates; the β-ADP site is converted to the β-ATP conformation, which promotes condensation of bound to form ATP; and the β-empty site becomes a β-ADP site, which loosely binds entering from the solvent. Note that the direction of rotation reverses when the ATP synthase is acting as an ATPase, as in the experiment depicted in Figure 19-27.
The conformational changes central to this mechanism are driven by the passage of protons through the portion of ATP synthase. The streaming of protons through the pore causes the c ring and the attached γ subunit to rotate about the long axis of γ, which is perpendicular to the plane of the membrane. The γ subunit passes through the center of the spheroid, which is held stationary relative to the membrane surface by the and subunits (Fig. 19-25a). With each rotation of 120°, γ comes into contact with a different β subunit, and the contact forces that β subunit into the β-empty conformation.
The three β subunits interact in such a way that when one assumes the β-empty conformation, its neighbor to one side must assume the β-ADP form, and the other neighbor the β-ATP form. Thus, one complete rotation of the γ subunit causes each β subunit to cycle through all three of its possible conformations, and for each rotation, three ATP are synthesized and released from the enzyme surface.
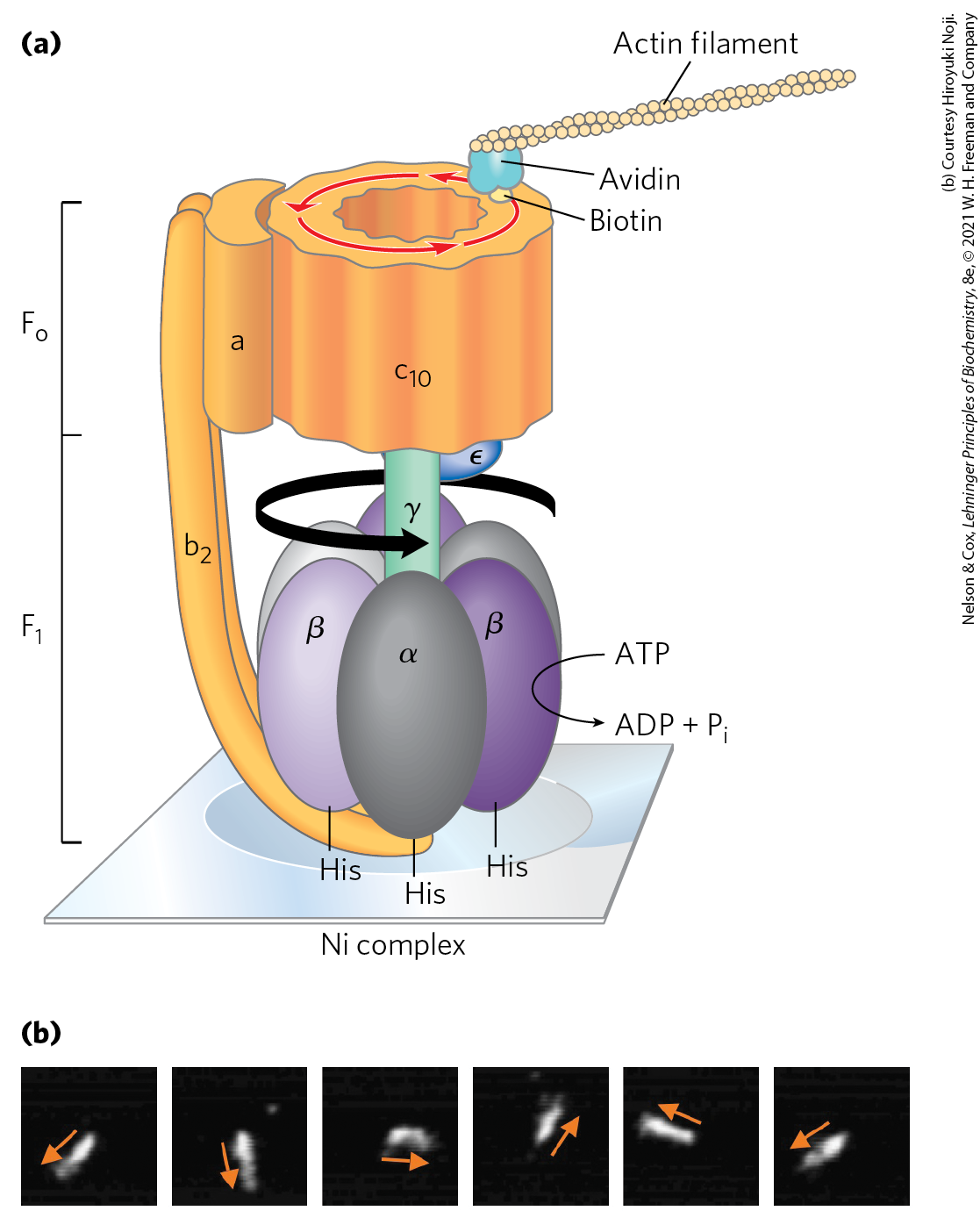
One strong prediction of this binding-change model is that the γ subunit should rotate in one direction when is synthesizing ATP and in the opposite direction when the enzyme is hydrolyzing ATP. This prediction of rotation with ATP hydrolysis was confirmed in elegant experiments in the laboratories of Masasuke Yoshida and Kazuhiko Kinosita, Jr. The rotation of γ in a single molecule was observed microscopically by attaching a long, thin, fluorescent actin polymer to γ and watching it move relative to immobilized on a microscope slide as ATP was hydrolyzed. (The expected reversal of the rotation when ATP is being synthesized could not be tested in this experiment; there is no proton gradient to drive ATP synthesis.) When the entire complex (not just ) was used in a similar experiment, the entire ring of c subunits rotated with γ (Fig. 19-27). The “shaft” rotated in the predicted direction through 360°. The rotation was not smooth but occurred in three discrete steps of 120°. As calculated from the known rate of ATP hydrolysis by one molecule and from the frictional drag on the long actin polymer, the efficiency of this mechanism in converting chemical energy into motion is close to 100%. It is, in Boyer’s words, “a splendid molecular machine!”
FIGURE 19-27 Experimental demonstration of rotation of and . This fundamental property of the ATP synthase reaction was demonstrated in several creative ways. (a) In one experiment, illustrated in this cartoon, , genetically engineered to contain a run of His residues, was tightly adhered to a microscope slide coated with a Ni complex; biotin was covalently attached to a c subunit of . The protein avidin, which binds biotin very tightly, was covalently attached to long filaments of actin labeled with a fluorescent probe. Biotin-avidin binding then attached the actin filaments to the c subunit. When ATP was provided as substrate for the ATPase activity of , the labeled filament rotated in one direction, proving that the cylinder of c subunits rotates. (b) In another experiment, a fluorescent actin filament was attached directly to the γ subunit. The series of fluorescence micrographs (read left to right) shows the position of the actin filament at intervals of 133 ms. Note that as the filament rotated, it made discrete jumps rather than smooth rotation about the circle. The cylinder and shaft move as one unit. [(a) Information from Y. Sambongi et al., Science 286:1722, 1999.]
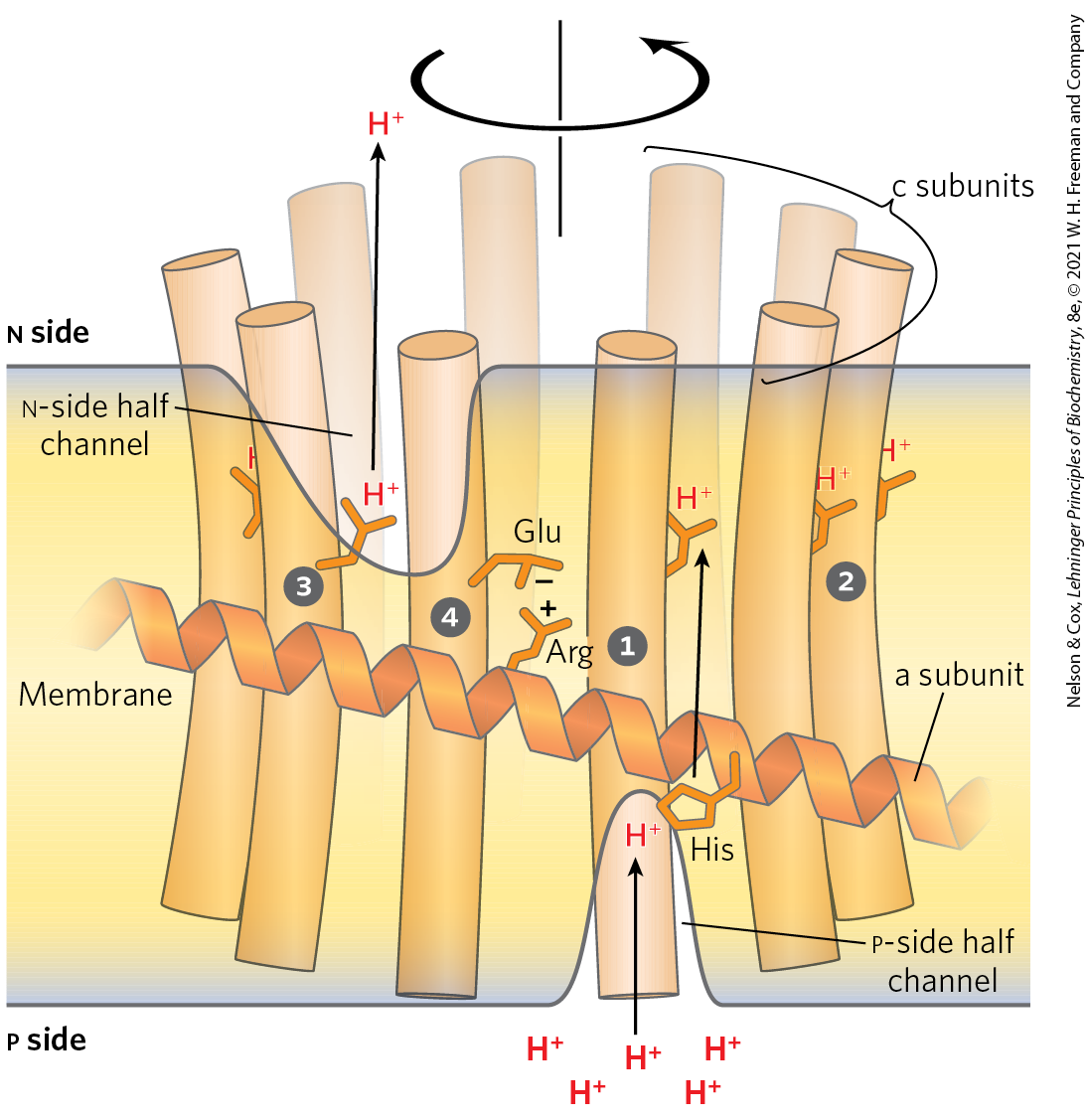
A model that illustrates how proton flow and rotary motion are coupled in the complex is shown in Figure 19-28. The a subunit is stationary, while the c ring rotates. Critical interactions occur between conserved amino acids in the a and c subunits. The individual subunits in the c ring are arranged in a circle with only a few in contact with the a subunit at any moment. Protons diffuse across the membrane through a path made up of both a and c subunits. Transient protonation of a key Glu residue in each c subunit elicits conformation changes that drive rotation and transmit protons between hydrophilic half channels positioned on each side of the membrane. The rotary movement of the c ring is made unidirectional by the large difference in proton concentration across the membrane. The number of protons that must be transferred to produce one complete rotation of the c ring is equal to the number of c subunits in the ring. Structural studies of the c ring have shown that the number of c subunits differs in different organisms (Fig. 19-29). In bovine mitochondria the number is 8, in yeast mitochondria and in Escherichia coli it is 10, and the number of c subunits can range as high as 17, as is seen in the soil bacterium Burkholderia pseudomallei. The rate of rotation in intact mitochondria has been estimated at about 6,000 rpm — 100 rotations per second.
FIGURE 19-28 A model for proton-driven rotation of the c ring. The a subunit of the complex of the ATP synthase (see Fig. 19-25a) has two hydrophilic half-channels for protons, one leading from the p side to the middle of the membrane, the other leading from the middle of the membrane to the n side (matrix). The function of the stationary a subunit is to conduct protons to and from the c ring subunits to drive c ring rotational motion. Individual c subunits in (the total number varies from 8 to 17 in different species) are arranged in a circle about a central core. Each c subunit has a critical Glu residue (an Asp in some species) about midway across the membrane, with a perturbed allowing it to donate or accept a proton (red ) at pH near neutrality. The cycle that c subunits go through is illustrated. One c subunit is initially positioned so that a proton that enters the half-channel on the p side (where the proton concentration is relatively high) encounters and protonates a conserved His residue in the a subunit, transferring it to the c subunit Glu residue. This triggers a rotation-facilitating conformational change in the protonated c subunit as the Glu loses its negative charge. The now neutral Glu residue is sequestered in the hydrophobic membrane layer as it rotates as part of the c ring . As the c ring rotates, the c subunit we are following eventually makes contact with the channel to the n side of the membrane, where the environment is relatively alkaline, and the proton is released . As the Glu reacquires its negative charge, another rotation-facilitating conformational change occurs such that the Glu interacts transiently with a conserved Arg residue in the immobile a subunit . The interaction with the Arg is disrupted as the Glu is again protonated by the His residue, in motions that again facilitate rotation. The c subunits positioned near the half channels are providing the rotational driving force at any given moment, as they are the ones undergoing conformation changes associated with protonation and deprotonation. The orientation of the proton gradient dictates the direction of proton flow and makes rotation of the c ring essentially unidirectional. [Information from W. Kühlbrandt and K. M. Davies, Trends Biochem. Sci. 41:106, 2016.]
Chemiosmotic Coupling Allows Nonintegral Stoichiometries of Consumption and ATP Synthesis
The overall reaction equation for ATP synthesis has the following form:
(19-11)
The value of x is sometimes called the P/O ratio or the . When a proton gradient is coupled to ATP synthesis as described above, there is no theoretical requirement for to be integral. The relevant questions about stoichiometry become these: How many protons are pumped outward by electron transfer from one NADH to ? and How many protons must flow inward through the complex to drive the synthesis of one ATP? The measurement of proton fluxes is technically complicated; the investigator must take into account the buffering capacity of mitochondria, nonproductive leakage of protons across the inner membrane, and use of the proton gradient for functions other than ATP synthesis, such as driving the transport of substrates across the inner mitochondrial membrane (described below). When NADH or succinate (which sends electrons into the respiratory chain at the level of ubiquinone) is the oxidizable substrate, the consensus experimental values for number of protons pumped out per pair of electrons are 10 and 6, respectively. The most widely accepted experimental value for number of protons required to drive the synthesis of an ATP molecule is 4, of which 1 is used in transporting , ATP, and ADP across the mitochondrial membrane (see below). If 10 protons are pumped out per NADH and 4 must flow in to produce 1 ATP, the proton-based ratio is 2.5 for NADH as the electron donor and for succinate. However, as we will see in Worked Example 19-2, the proton stoichiometry of ATP synthesis by ATP synthase depends upon the number of c units in , which ranges from 8 to 17, depending on the species.
WORKED EXAMPLE 19-2 Stoichiometry of ATP Production: Effect of c Ring Size
(a) If the ATP synthase of bovine mitochondria has 8 c subunits per c ring, what is the predicted ratio of ATP formed per NADH oxidized? (b) What is the predicted value for yeast mitochondria, with 10 c subunits per ATP synthase? (c) What are the comparable values for electrons entering the respiratory chain from ?
SOLUTION:
(a) Here we are asked to determine how many ATP molecules are produced per NADH. This is another way of asking us to calculate the ratio, or x, in Equation 19-11. If the c ring has 8 c subunits, then one full rotation will transfer 8 protons to the matrix and produce 3 ATP molecules. But this synthesis also requires the transport of 3 into the matrix, at a cost of 1 proton each, adding 3 more protons to the total number required. This brings the total cost to . The generally agreed value for the number of protons pumped out per pair of electrons transferred from NADH is 10 (Eqn 19-7). So, oxidizing 1 NADH produces .
(b) If the c ring has 10 c subunits, then one full rotation will transfer 10 protons to the matrix and produce 3 ATP molecules. Adding in the 3 protons to transport the 3 into the matrix brings the total cost to . Oxidizing 1 NADH produces .
(c) When electrons enter the respiratory chain from (at ubiquinone), only 6 protons are available to drive ATP synthesis. This changes the calculation for bovine mitochondria to per pair of electrons from . For yeast mitochondria, the calculation is per pair of electrons from .
These calculated values of x, or the ratio, define a range that includes the experimental values of and , and we therefore use these values throughout this book.
The Proton-Motive Force Energizes Active Transport
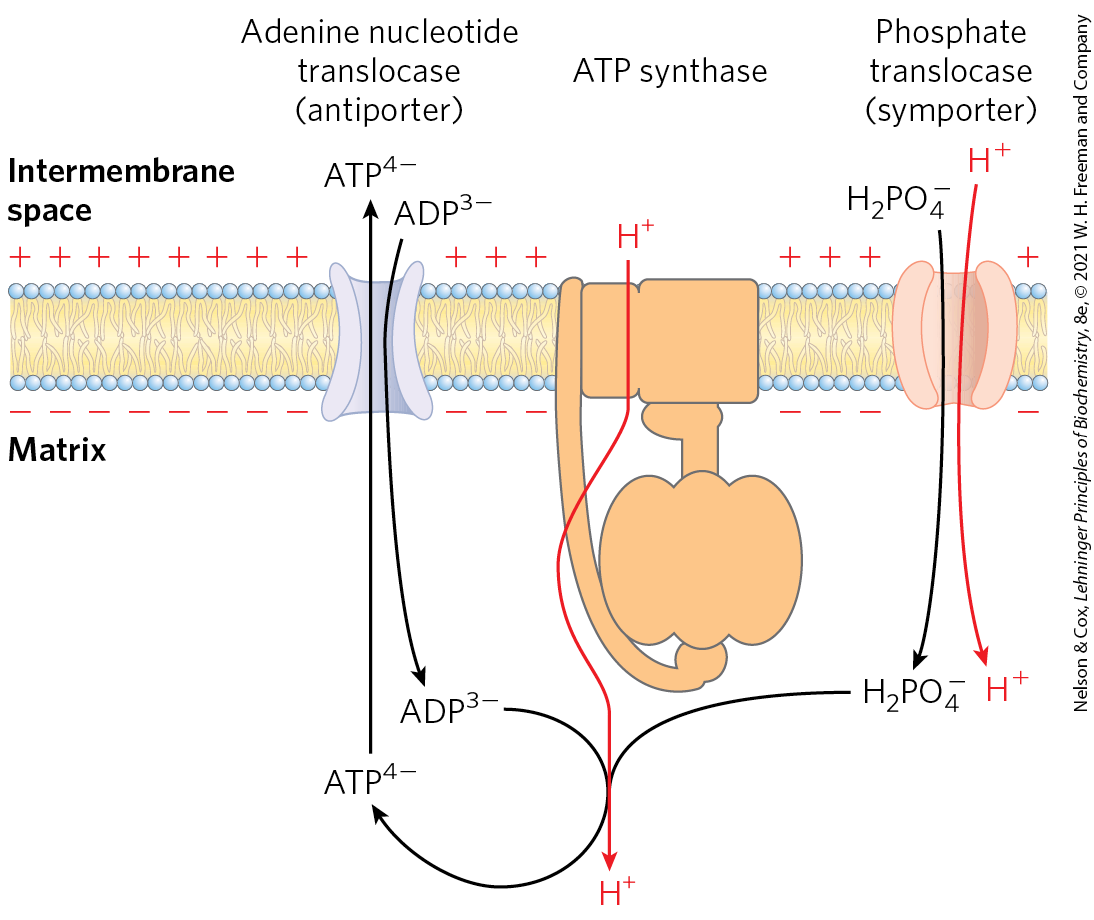
Although the primary role of the proton gradient in mitochondria is to furnish energy for the synthesis of ATP, the proton-motive force also drives several transport processes essential to oxidative phosphorylation. The inner mitochondrial membrane is generally impermeable to charged species, but two specific systems transport ADP and into the matrix and ATP out to the cytosol (Fig. 19-30).
FIGURE 19-30 Adenine nucleotide and phosphate translocases. Transport systems of the inner mitochondrial membrane carry ADP and into the matrix and newly synthesized ATP into the cytosol. The adenine nucleotide translocase is an antiporter; the same protein moves ADP into the matrix and ATP out. The effect of replacing with in the matrix is the net efflux of one negative charge, which is favored by the charge difference across the inner membrane (outside positive). At pH 7, is present as both and ; the phosphate translocase is specific for . There is no net flow of charge during symport of and , but the relatively low proton concentration in the matrix favors the inward movement of . Thus the proton-motive force is responsible both for providing the energy for ATP synthesis and for transporting substrates (ADP and ) into and product (ATP) out of the mitochondrial matrix. All three of these transport systems can be isolated as a single membrane-bound complex (ATP synthasome).
The adenine nucleotide translocase, integral to the inner membrane, binds in the intermembrane space and transports it into the matrix in exchange for an molecule simultaneously transported outward (see Fig. 13-11 for the ionic forms of ATP and ADP). Because this antiporter moves four negative charges out for every three moved in, its activity is favored by the transmembrane electrochemical gradient, which gives the matrix a net negative charge; the proton-motive force drives ATP-ADP exchange. Adenine nucleotide translocase is specifically inhibited by atractyloside, a toxic glycoside produced by a species of thistle. If the transport of ADP into and ATP out of mitochondria is inhibited, cytosolic ATP cannot be regenerated from ADP, explaining the toxicity of atractyloside.
A second membrane transport system essential to oxidative phosphorylation is the phosphate translocase, which promotes symport of one and one into the matrix. This transport process, too, is favored by the transmembrane proton gradient (Fig. 19-30). Notice that the process requires movement of one proton from the p side to the n side of the inner membrane, consuming some of the energy of electron transfer. A complex of the ATP synthase and both translocases, the ATP synthasome, can be isolated from mitochondria by gentle dissection with detergents, suggesting that the functions of these three proteins are very tightly integrated.
ATP and ADP cross the outer mitochondrial membrane via the voltage-dependent anion channel (VDAC), a 19-stranded β barrel with an opening about 27 Å wide, connecting the cytosol and the intermembrane space. Each VDAC, when open, can move ATP molecules per second. The opening is gated by voltage, as its name indicates, and under some conditions VDAC is closed to ATP.
Shuttle Systems Indirectly Convey Cytosolic NADH into Mitochondria for Oxidation
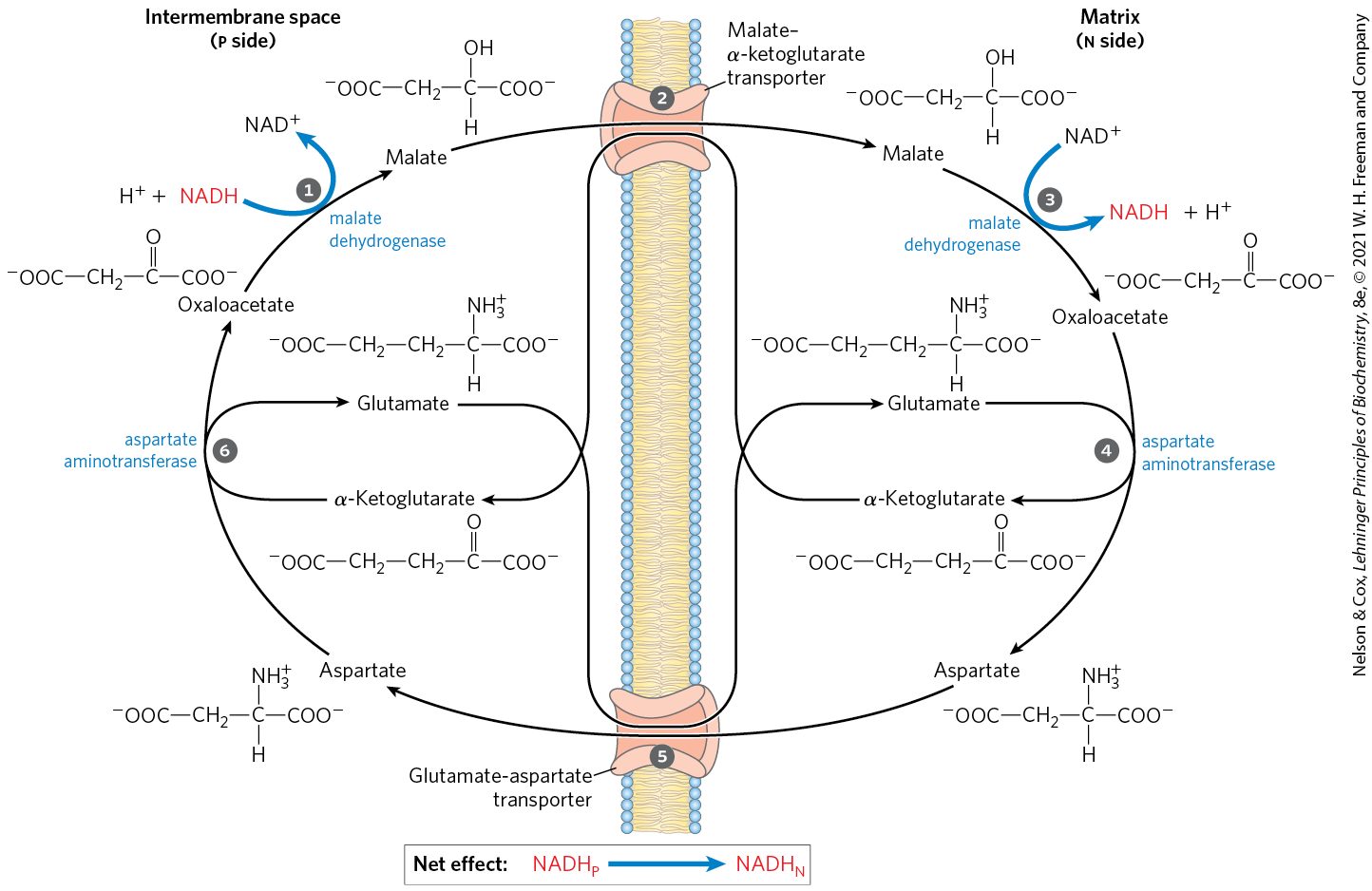
The NADH dehydrogenase of the inner mitochondrial membrane of animal cells can accept electrons only from NADH in the matrix. Given that the inner membrane is not permeable to NADH, how can the NADH generated by glycolysis in the cytosol be reoxidized to by via the respiratory chain? Special shuttle systems carry reducing equivalents from cytosolic NADH into mitochondria by an indirect route. The most active NADH shuttle, which functions in liver, kidney, and heart mitochondria, is the malate-aspartate shuttle (Fig. 19-31). The reducing equivalents of cytosolic NADH are first transferred to cytosolic oxaloacetate to yield malate, catalyzed by cytosolic malate dehydrogenase. The malate thus formed passes through the inner membrane via the malate–α-ketoglutarate transporter. Within the matrix, the reducing equivalents are passed to by the action of matrix malate dehydrogenase, forming NADH; this NADH can pass electrons directly to the respiratory chain. About 2.5 molecules of ATP are generated as this pair of electrons passes to . Cytosolic oxaloacetate must be regenerated by transamination reactions and the activity of membrane transporters to start another cycle of the shuttle.
FIGURE 19-31 Malate-aspartate shuttle. This shuttle for transporting reducing equivalents from cytosolic NADH into the mitochondrial matrix is used in liver, kidney, and heart. NADH in the cytosol enters the intermembrane space through openings in the outer membrane (porins), then passes two reducing equivalents to oxaloacetate, producing malate. Malate crosses the inner membrane via the malate–α-ketoglutarate transporter. In the matrix, malate passes two reducing equivalents to , and the resulting NADH is oxidized by the respiratory chain; the oxaloacetate formed from malate cannot pass directly into the cytosol. Oxaloacetate is first transaminated to aspartate, and aspartate can leave via the glutamate-aspartate transporter. Oxaloacetate is regenerated in the cytosol, completing the cycle, and glutamate produced in the same reaction enters the matrix via the glutamate-aspartate transporter.
Skeletal muscle and brain use a different NADH shuttle, the glycerol 3-phosphate shuttle (Fig. 19-32). It differs from the malate-aspartate shuttle in that it delivers the reducing equivalents from NADH through FAD in glycerol 3-phosphate dehydrogenase to ubiquinone and thus into Complex III, not Complex I (Fig. 19-15), providing enough energy to synthesize only 1.5 ATP molecules per pair of electrons.
FIGURE 19-32 Glycerol 3-phosphate shuttle. This alternative means of moving reducing equivalents from the cytosol to the respiratory chain operates in skeletal muscle and the brain. In the cytosol, dihydroxyacetone phosphate accepts two reducing equivalents from NADH in a reaction catalyzed by cytosolic glycerol 3-phosphate dehydrogenase. An isozyme of glycerol 3-phosphate dehydrogenase bound to the outer face of the inner membrane then transfers two reducing equivalents from glycerol 3-phosphate in the intermembrane space to ubiquinone. Note that this shuttle does not involve membrane transport systems.
The mitochondria of plants have an externally oriented NADH dehydrogenase that can transfer electrons directly from cytosolic NADH into the respiratory chain at the level of ubiquinone. Because this pathway bypasses the NADH dehydrogenase of Complex I and the associated proton movement, the yield of ATP from cytosolic NADH is less than that from NADH generated in the matrix (Box 19-1).
SUMMARY 19.2 ATP Synthesis
The chemiosmotic theory describes the coupling of ATP synthesis to an electrochemical proton gradient. The flow of electrons through Complexes I, III, and IV results in pumping of protons across the inner mitochondrial membrane, making the matrix alkaline relative to the intermembrane space. This proton gradient provides the energy, in the form of the proton-motive force, for ATP synthesis from ADP and .
ATP synthase has two major components, called and . Both components have multiple subunits. The overall complex spans the inner mitochondrial membrane.
ATP synthesis is reversible within the active site on the β subunits of the complex. Very tight binding to ATP offsets the negative for ATP hydrolysis in solution.
Release of ATP from ATP synthase is promoted by the transmembrane proton gradient.
The subunits of the complex cycle from –bound to ATP-bound to empty conformations.
ATP synthase carries out “rotational catalysis,” in which the flow of protons through causes the c ring to rotate and in turn trigger the subunit conformational changes in .
The ratio of ATP synthesized per reduced to (the ratio) is about 2.5 when electrons enter the respiratory chain at Complex I, and 1.5 when electrons enter at ubiquinone. This ratio varies among species, depending on the number of c subunits in the complex.
Energy conserved in a proton gradient can drive solute transport uphill across a membrane.
The inner mitochondrial membrane is impermeable to NADH and , but NADH equivalents are moved from the cytosol to the matrix by either of two shuttles. NADH equivalents moved in by the malate-aspartate shuttle enter the respiratory chain at Complex I and yield a ratio of 2.5; those moved in by the glycerol 3-phosphate shuttle enter at ubiquinone and give a ratio of 1.5.

 When isolated mitochondria are suspended in a buffer containing ADP, , and an oxidizable substrate such as succinate, three easily measured processes occur: (1) the substrate is oxidized (succinate yields fumarate), (2) is consumed, and (3) ATP is synthesized. Oxygen consumption and ATP synthesis depend on the presence of an oxidizable substrate (succinate in this case) as well as ADP and .
When isolated mitochondria are suspended in a buffer containing ADP, , and an oxidizable substrate such as succinate, three easily measured processes occur: (1) the substrate is oxidized (succinate yields fumarate), (2) is consumed, and (3) ATP is synthesized. Oxygen consumption and ATP synthesis depend on the presence of an oxidizable substrate (succinate in this case) as well as ADP and .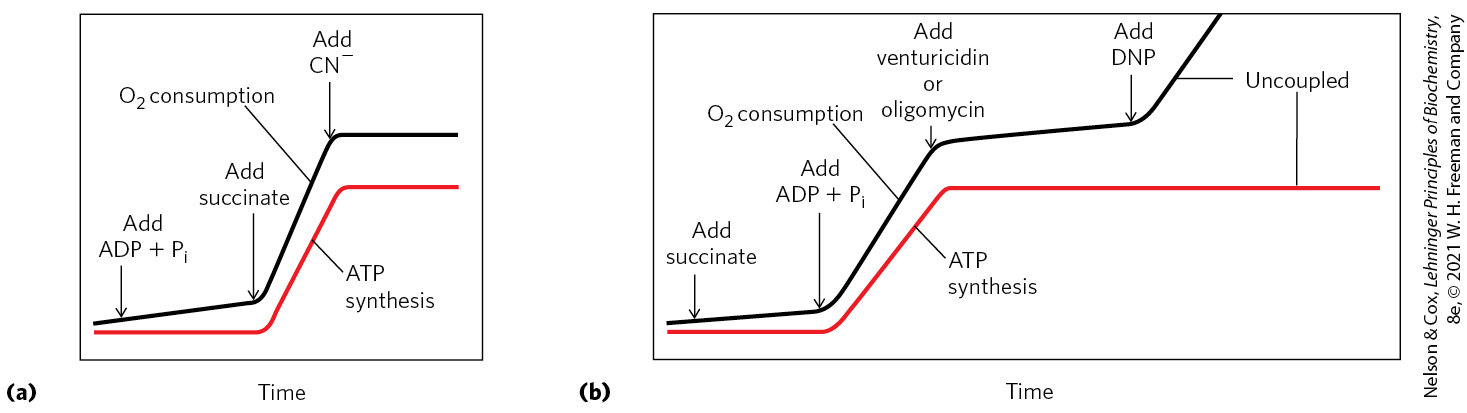

 Mitochondrial ATP synthase is an F-type ATPase (see
Mitochondrial ATP synthase is an F-type ATPase (see 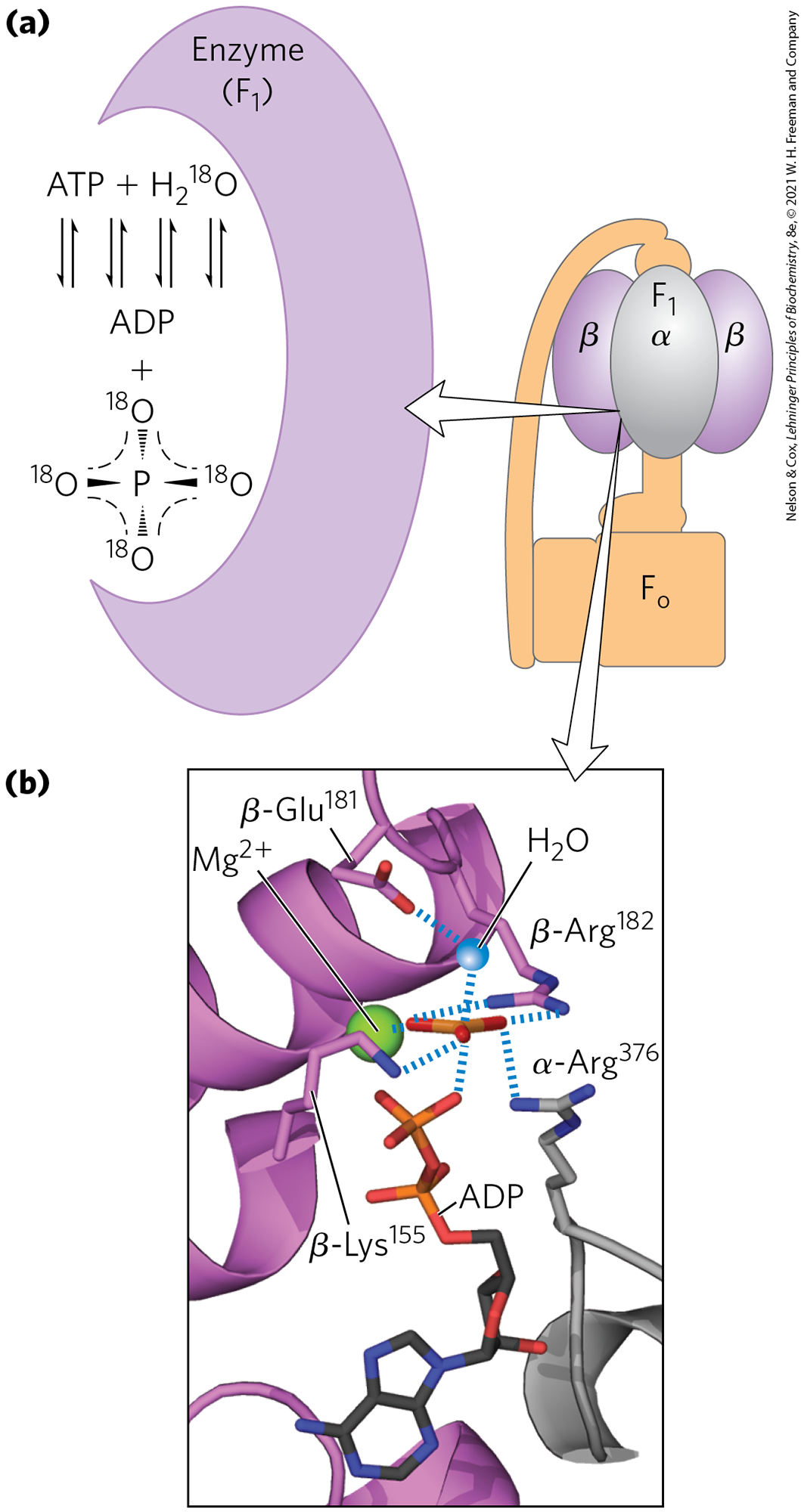
 is initially positioned so that a proton that enters the half-channel on the
is initially positioned so that a proton that enters the half-channel on the  . As the c ring rotates, the c subunit we are following eventually makes contact with the channel to the
. As the c ring rotates, the c subunit we are following eventually makes contact with the channel to the  . As the Glu reacquires its negative charge, another rotation-facilitating conformational change occurs such that the Glu interacts transiently with a conserved Arg residue in the immobile a subunit
. As the Glu reacquires its negative charge, another rotation-facilitating conformational change occurs such that the Glu interacts transiently with a conserved Arg residue in the immobile a subunit  . The interaction with the Arg is disrupted as the Glu is again protonated by the His residue, in motions that again facilitate rotation. The c subunits positioned near the half channels are providing the rotational driving force at any given moment, as they are the ones undergoing conformation changes associated with protonation and deprotonation. The orientation of the proton gradient dictates the direction of proton flow and makes rotation of the c ring essentially unidirectional.
. The interaction with the Arg is disrupted as the Glu is again protonated by the His residue, in motions that again facilitate rotation. The c subunits positioned near the half channels are providing the rotational driving force at any given moment, as they are the ones undergoing conformation changes associated with protonation and deprotonation. The orientation of the proton gradient dictates the direction of proton flow and makes rotation of the c ring essentially unidirectional. 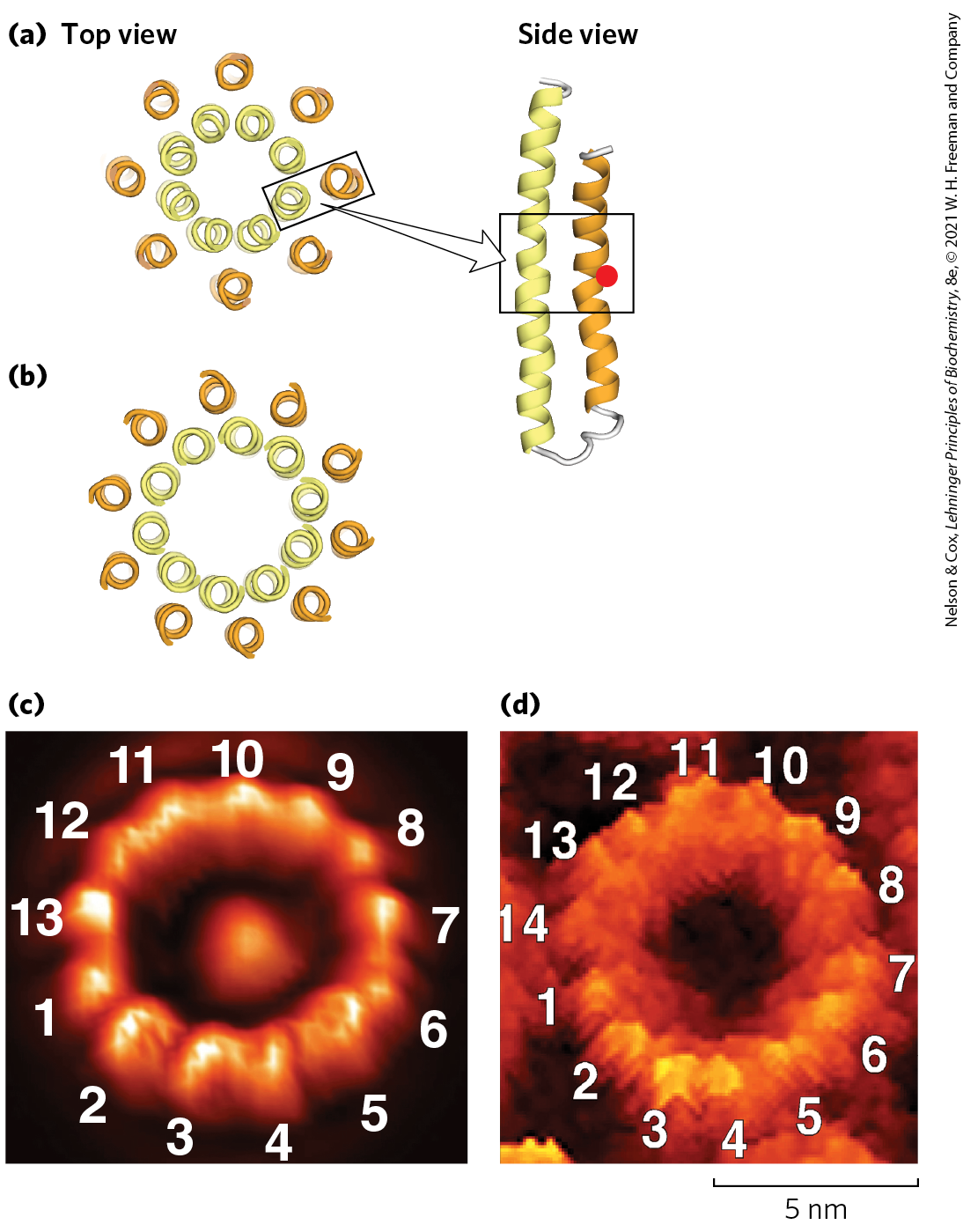
 aspartate can leave via the glutamate-aspartate transporter.
aspartate can leave via the glutamate-aspartate transporter.  Oxaloacetate is regenerated in the cytosol, completing the cycle, and glutamate produced in the same reaction enters the matrix via the glutamate-aspartate transporter.
Oxaloacetate is regenerated in the cytosol, completing the cycle, and glutamate produced in the same reaction enters the matrix via the glutamate-aspartate transporter.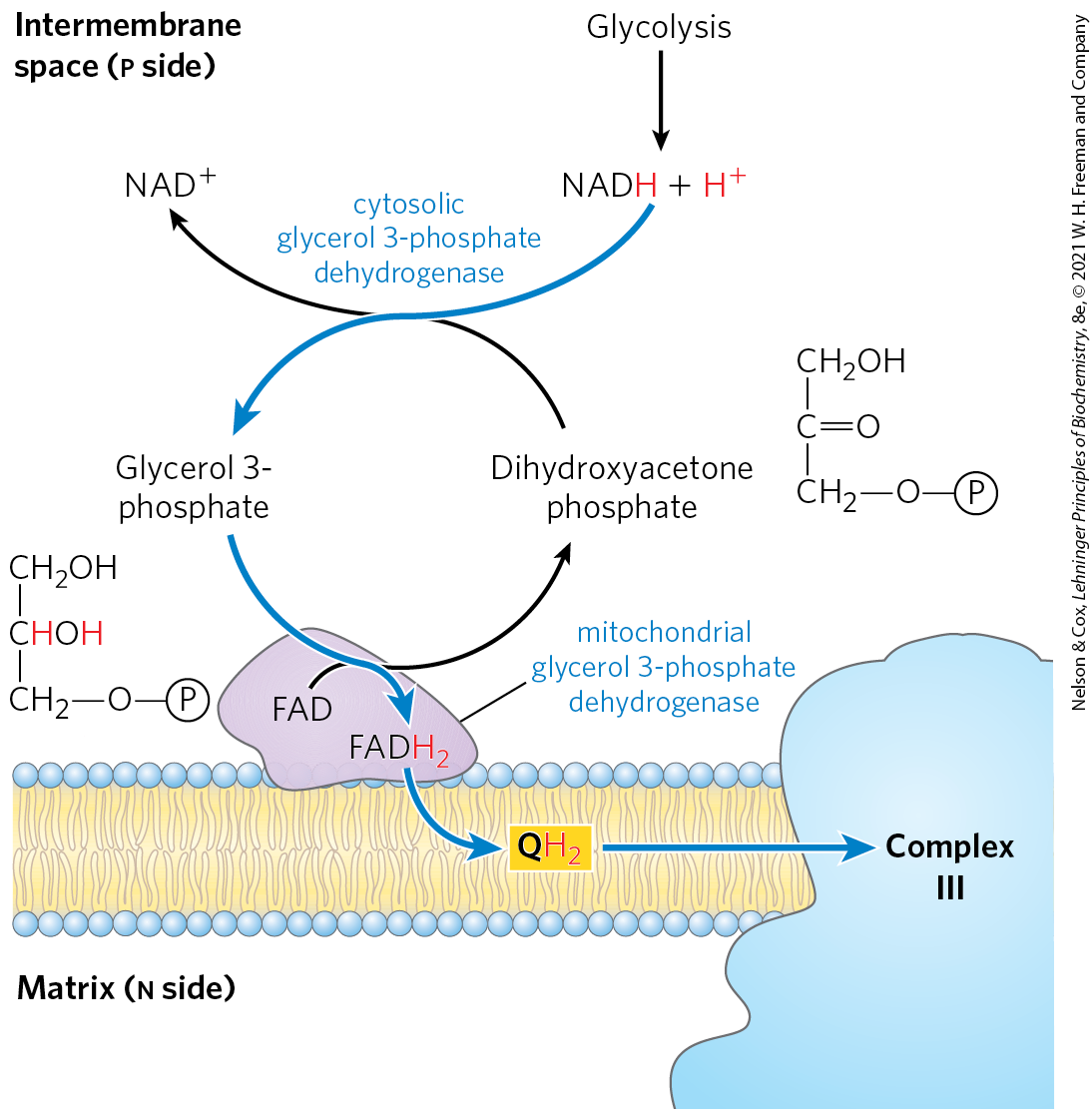
 The chemiosmotic theory describes the coupling of ATP synthesis to an electrochemical proton gradient. The flow of electrons through Complexes I, III, and IV results in pumping of protons across the inner mitochondrial membrane, making the matrix alkaline relative to the intermembrane space. This proton gradient provides the energy, in the form of the proton-motive force, for ATP synthesis from ADP and .
The chemiosmotic theory describes the coupling of ATP synthesis to an electrochemical proton gradient. The flow of electrons through Complexes I, III, and IV results in pumping of protons across the inner mitochondrial membrane, making the matrix alkaline relative to the intermembrane space. This proton gradient provides the energy, in the form of the proton-motive force, for ATP synthesis from ADP and .