Chapter Review
KEY TERMS
Terms in bold are defined in the glossary.
- photosynthesis
- light-dependent reactions
- photophosphorylation
- chloroplast
- stroma
- thylakoid
- photon
- excited state
- ground state
- exciton
- exciton transfer
- chlorophylls
- accessory pigments
- carotenoids
- β-carotene
- action spectrum
- photosystem
- photochemical reaction center
- light-harvesting complexes (LHCs)
- cyclic electron transfer
- linear electron transfer
- ferredoxin
- Z scheme
- photosystem II (PSII)
- photosystem I (PSI)
- cytochrome
- plastoquinone
- plastocyanin
- phylloquinone
- cyclic photophosphorylation
- state transition
- oxygen-evolving center
- assimilation
- fixation
- Calvin cycle
- reductive pentose phosphate pathway
- ribulose 1,5-bisphosphate
- 3-phosphoglycerate
- plants ribulose 1,5-bisphosphate carboxylase/oxygenase (rubisco)
- rubisco activase
- thioredoxin
- ferredoxin:thioredoxin reductase
- photorespiration
- 2-phosphoglycolate
- glycolate pathway
- plants
- pathway
- phosphoenolpyruvate carboxylase
- malic enzyme
- pyruvate phosphate dikinase
- CAM plants
- sugar nucleotide
- starch synthase
- glyoxysome
- glyoxylate cycle
- isocitrate lyase
- glyoxylate
- malate synthase
- cellulose synthase
Problems
1. Photochemical Efficiency of Light at Different Wavelengths The rate of photosynthesis in a green plant, measured by production, is higher when illuminated with light of wavelength 680 nm than with light of wavelength 700 nm. However, illumination by a combination of light of 680 nm and 700 nm gives a higher rate of photosynthesis than light of either wavelength alone. Explain.
2. Balance Sheet for Photosynthesis In 1804, Nicolas-Théodore de Saussure observed that the total weight of oxygen and dry organic matter produced by plants is greater than the weight of carbon dioxide consumed during photosynthesis. Where does the extra weight come from?
3. Role of in Some Photosynthetic Bacteria Illuminated purple sulfur bacteria carry out photosynthesis in the presence of and , but only if is added and is absent. During photosynthesis, measured by formation of carbohydrate, the bacteria convert to elemental sulfur but do not produce . What is the role of the conversion of to sulfur? Why doesn’t photosynthesis produce in these bacteria?
4. Electron Transfer through Photosystems I and II Predict how an inhibitor of electron passage through pheophytin would affect electron transfer through (a) photosystem II and (b) photosystem I. Explain your reasoning.
5. Limited ATP Synthesis in the Dark In a laboratory experiment, a researcher illuminates spinach chloroplasts in the absence of ADP and . Then, the researcher turns the light off and adds ADP and . ATP synthesis occurs for a short time in the dark. Explain this finding.
6. Mode of Action of the Herbicide DCMU Treating chloroplasts with 3-(3,4-dichlorophenyl)-1,1-dimethylurea (DCMU, or diuron), a potent herbicide, causes oxygen evolution and photophosphorylation to cease. Adding a Hill reagent (an external electron acceptor) restores oxygen evolution, but not photophosphorylation. How does DCMU act as a weed killer? Suggest a location for the inhibitory action of this herbicide in the scheme shown in Figure 20-12. Explain.
7. Effect of Venturicidin on Oxygen Evolution Venturicidin is a powerful inhibitor of the chloroplast ATP synthase, interacting with and blocking proton passage through the complex. How would venturicidin affect oxygen evolution in a suspension of well-illuminated chloroplasts? Would your answer change if the experiment were done in the presence of an uncoupling reagent such as 2,4-dinitrophenol (DNP)? Explain.
8. Light Energy for a Redox Reaction Suppose you have isolated a new photosynthetic microorganism that oxidizes and passes the electrons to . What wavelength of light would provide enough energy for to reduce under standard conditions? Assume 100% efficiency in the photochemical event, and use an of for and for . See Figure 20-4 for the energy equivalents of wavelengths of light.
9. Equilibrium Constant for Water-Splitting Reactions The coenzyme is the terminal electron acceptor in chloroplasts, according to the reaction
Use information in Chapter 19 (Table 19-2) to calculate the equilibrium constant for this reaction at 25 °C. (The relationship between and is discussed on p. 468.) How can the chloroplast overcome this unfavorable equilibrium?
10. Energetics of Phototransduction During photosynthesis, pigment molecules in chloroplasts must absorb eight photons (four by each photosystem) for every molecule they produce, according to the equation
The for the light-independent production of is 400 kJ/mol. Assuming that these photons have a wavelength of 700 nm (red) and that the light absorption and use of light energy are 100% efficient, calculate the free-energy change for the process.
11. Electron Transfer to a Hill Reagent Isolated spinach chloroplasts evolve when illuminated in the presence of potassium ferricyanide (a Hill reagent), according to the equation
where represents ferricyanide and represents ferrocyanide. Does this process produce NADPH? Explain.
12. How Often Does a Chlorophyll Molecule Absorb a Photon? The amount of chlorophyll a in a spinach leaf is about of leaf surface. In noonday sunlight (average energy reaching the leaf is ), the leaf absorbs about 50% of the radiation. How often does a single chlorophyll molecule absorb a photon? Given that the average lifetime of an excited chlorophyll molecule in vivo is 1 ns, what fraction of the chlorophyll molecules are excited at any one time?
13. Effect of Monochromatic Light on Electron Flow Using a spectrophotometer, researchers can sometimes directly observe the extent of oxidation or reduction of an electron carrier during photosynthetic electron transfer. Illuminating chloroplasts with 700 nm light oxidizes cytochrome f, plastocyanin, and plastoquinone. Illuminating chloroplasts with 680 nm light, however, reduces these electron carriers. Explain.
14. Function of Cyclic Photophosphorylation When the ratio in chloroplasts is high, photophosphorylation is predominantly cyclic (see Fig. 20-12). Does cyclic electron transfer evolve ? Does cyclic electron transfer produce NADPH? Explain. What is the main function of cyclic electron transfer?
15. Phases of Photosynthesis A researcher illuminates a suspension of green algae in the absence of . He then incubates the algae with in the dark and observes the conversion of to glucose for a brief time. What is the significance of this observation with regard to the -assimilation process, and how is it related to the light-dependent reactions of photosynthesis? Why does the conversion of to glucose stop after a brief time?
16. Identification of Key Intermediates in Assimilation Calvin and his colleagues used the unicellular green alga Chlorella to study the -assimilation reactions of photosynthesis. They incubated with illuminated suspensions of algae and followed the time course of appearance of in two compounds, X and Y, under two sets of conditions. Suggest the identities of X and Y, based on your understanding of the Calvin cycle.
- They grew illuminated Chlorella with unlabeled , then turned off the light and added (vertical dashed line in the graph below). Under these conditions, X was the first compound to become labeled with ; Y was unlabeled.
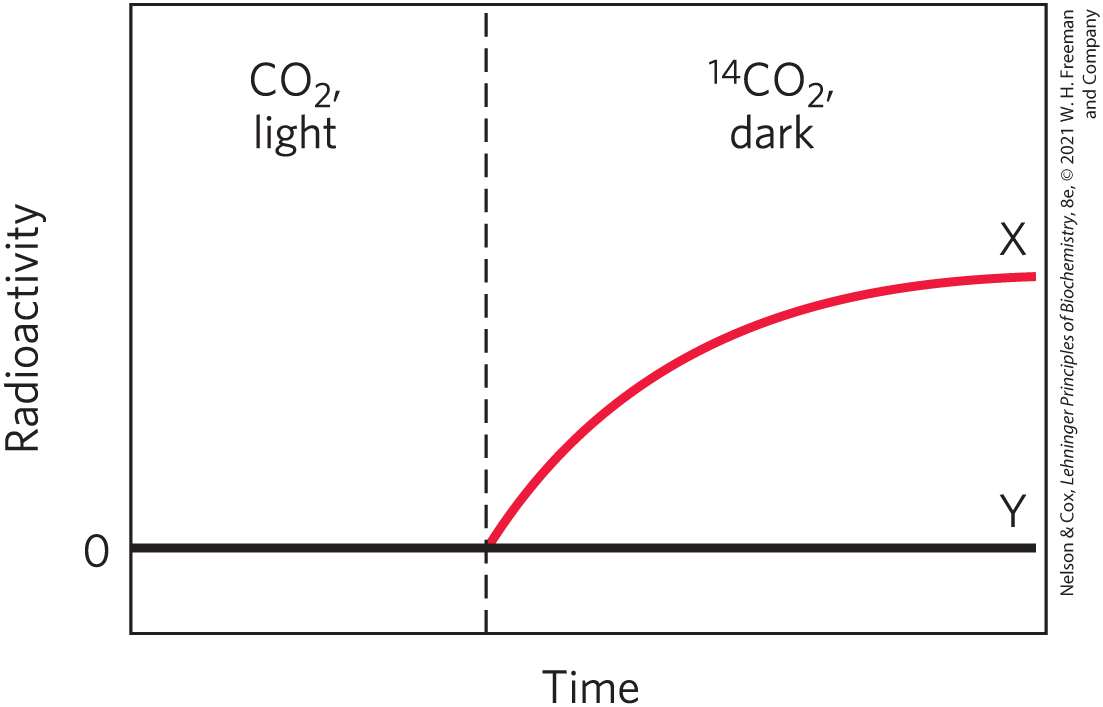
- They grew illuminated Chlorella cells with . Illumination was continued until all the had been taken up (vertical dashed line in the graph below). Under these conditions, X became labeled quickly but lost its radioactivity with time, whereas Y became more radioactive with time.
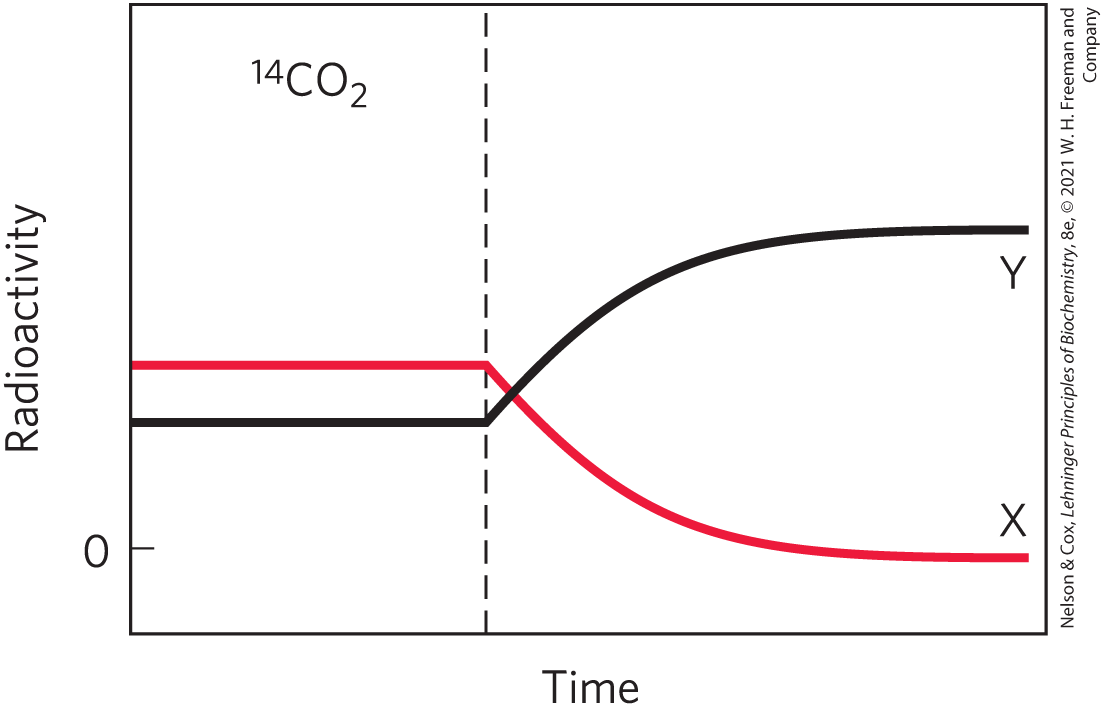
- They grew illuminated Chlorella with unlabeled , then turned off the light and added (vertical dashed line in the graph below). Under these conditions, X was the first compound to become labeled with ; Y was unlabeled.
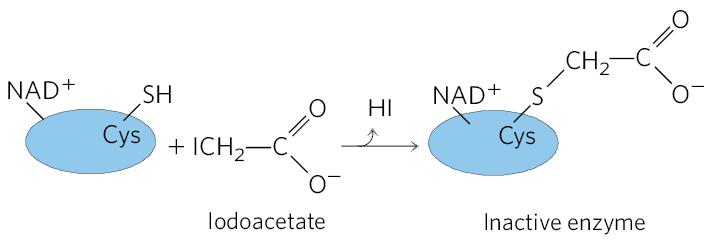
17. Regulation of the Calvin Cycle Iodoacetate reacts irreversibly with the free — SH groups of Cys residues in proteins. Predict which Calvin cycle enzyme(s) would be inhibited by iodoacetate, and explain why.
18. Comparison of the Reductive and Oxidative Pentose Phosphate Pathways The reductive pentose phosphate pathway generates several intermediates identical to those of the oxidative pentose phosphate pathway (Chapter 14). What role does each pathway play in cells where it is active?
19. Photorespiration and Mitochondrial Respiration Compare the oxidative photosynthetic carbon cycle, also called photorespiration, with the mitochondrial respiration that drives ATP synthesis. Why are both processes referred to as respiration? Where in the cell do they occur, and under what circumstances? What is the path of electron flow in each?
20. Pathway of Assimilation in Maize Researchers illuminate a maize (corn) plant in the presence of . After about 1 second of illumination, they find more than 90% of all the radioactivity incorporated in the leaves at C-4 of malate, aspartate, and oxaloacetate. Only after 60 seconds does appear at C-1 of 3-phosphoglycerate. Explain.
21. Identifying CAM Plants Given some and all the tools typically present in a biochemistry research lab, how would you design a simple experiment to determine whether a plant is a typical plant or a CAM plant?
22. Chemistry of Malic Enzyme: Variation on a Theme Malic enzyme, found in the bundle-sheath cells of plants, carries out a reaction that has a counterpart in the citric acid cycle. What is the analogous reaction? Explain your choice.
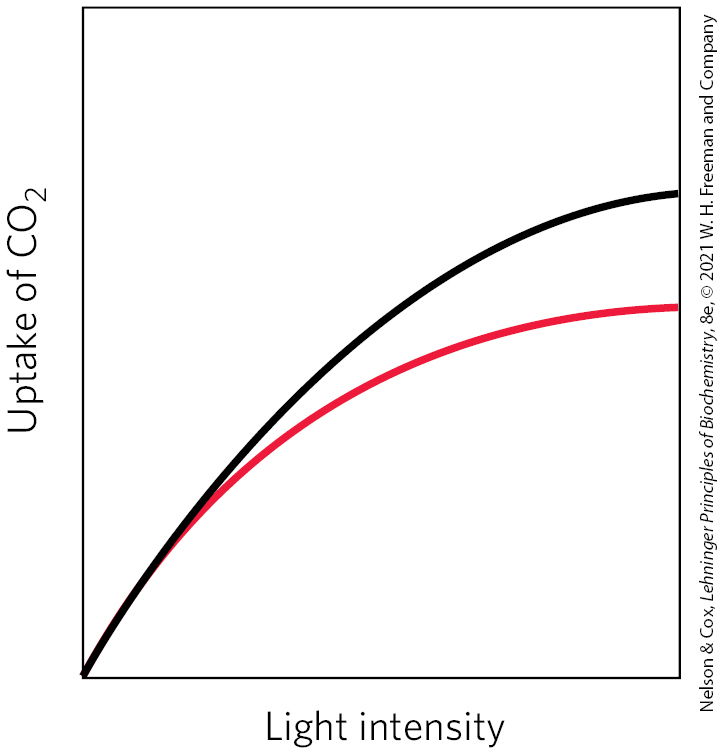
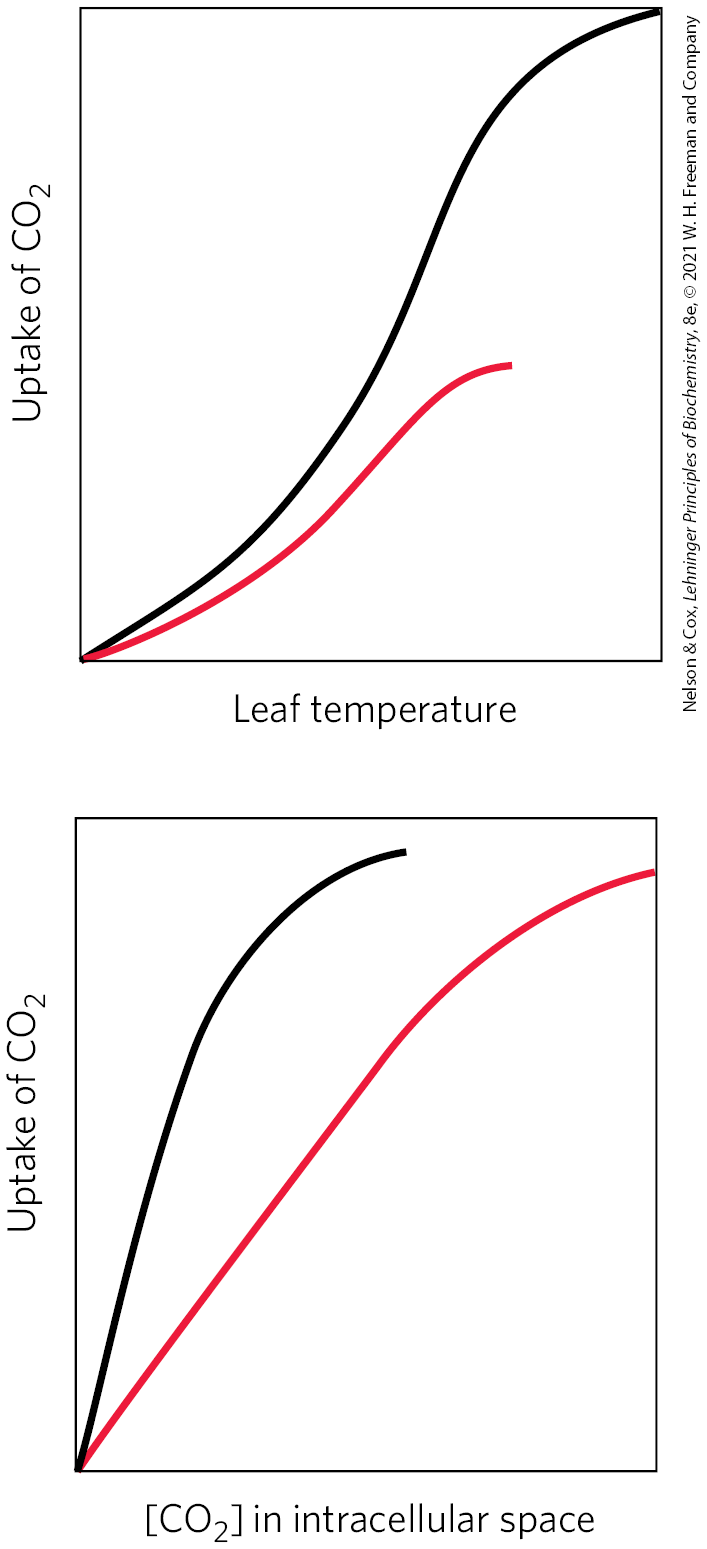
23. Differences between and Plants The plant genus Atriplex includes some and some species. In the plots, the black curve represents species 1; the red curve represents species 2. From the data in the plots, identify which is a plant and which is a plant. Justify your answer in molecular terms that account for the data in all three plots.
24. Inorganic Pyrophosphatase The enzyme inorganic pyrophosphatase contributes to making many biosynthetic reactions that generate inorganic pyrophosphate essentially irreversible in cells. By keeping the concentration of very low, the enzyme “pulls” these reactions in the direction of formation. The synthesis of ADP-glucose in chloroplasts is one such reaction. However, the synthesis of UDP-glucose in the plant cytosol, which also produces , is readily reversible in vivo. How do you reconcile these two facts?
25. Regulation of Starch and Sucrose Synthesis Sucrose synthesis occurs in the cytosol and starch synthesis occurs in the chloroplast stroma, yet the two processes are intricately balanced. What factors shift the reactions in favor of (a) starch synthesis and (b) sucrose synthesis?
26. Regulation of Sucrose Synthesis In the regulation of sucrose synthesis from the triose phosphates produced during photosynthesis, 3-phosphoglycerate and play critical roles (see Fig. 20-42). Explain why the concentrations of these two regulators reflect the rate of photosynthesis.
27. Sucrose and Dental Caries The most prevalent infection in humans worldwide is dental caries, which stems from the colonization and destruction of tooth enamel by a variety of acidifying microorganisms. These organisms synthesize and live within a water-insoluble network of dextrans, called dental plaque, composed of -linked polymers of glucose with many branch points. Polymerization of dextran requires dietary sucrose, and the bacterial enzyme dextran-sucrose glucosyltransferase catalyzes the reaction.
- Write the overall reaction for dextran polymerization.
- In addition to providing a substrate for the formation of dental plaque, how does dietary sucrose also provide oral bacteria with an abundant source of metabolic energy?
28. Partitioning between the Citric Acid and Glyoxylate Cycles In an organism (such as Escherichia coli) that has both the citric acid cycle and the glyoxylate cycle, what determines which of these pathways isocitrate will enter?
DATA ANALYSIS PROBLEM
29. Photophosphorylation: Discovery, Rejection, and Rediscovery In the 1930s and 1940s, researchers were beginning to make progress toward understanding the mechanism of photosynthesis. At the time, the role of “energy-rich phosphate bonds” (today, “ATP”) in glycolysis and cellular respiration was just becoming known. There were many theories about the mechanism of photosynthesis, especially about the role of light. This problem focuses on what was then called the “primary photochemical process” — that is, on what, exactly, the energy from captured light produces in the photosynthetic cell. Interestingly, one important part of the modern model of photosynthesis was proposed early on, only to be rejected, ignored for several years, then finally revived and accepted.
In 1944, Emerson, Stauffer, and Umbreit proposed that “the function of light energy in photosynthesis is the formation of ‘energy-rich’ phosphate bonds” (p. 107). In their model (hereafter, the “Emerson model”), the free energy necessary to drive both fixation and reduction came from these “energy-rich phosphate bonds” (i.e., ATP), produced as a result of light absorption by a chlorophyll-containing protein.
This model was explicitly rejected by Rabinowitch (1945). After summarizing Emerson and coauthors’ findings, Rabinowitch stated: “Until more positive evidence is provided, we are inclined to consider as more convincing a general argument against this hypothesis, which can be derived from energy considerations. Photosynthesis is eminently a problem of energy accumulation. What good can be served, then, by converting light quanta (even those of red light, which amount to about 43 kcal per Einstein) into ‘phosphate quanta’ of only 10 kcal per mole? This appears to be a start in the wrong direction — toward dissipation rather than toward accumulation of energy” (p. 228). This argument, along with other evidence, led to abandonment of the Emerson model until the 1950s, when it was found to be correct — albeit in a modified form.
For each piece of information from Emerson and coauthors’ article presented in (a) through (d), answer the following three questions:
How does this information support the Emerson model, in which light energy is used directly by chlorophyll to make ATP, and the ATP then provides the energy to drive fixation and reduction?
How would Rabinowitch explain this information, based on his model (and most other models of the day), in which light energy is used directly by chlorophyll to make reducing compounds? Rabinowitch wrote: “Theoretically, there is no reason why all electronic energy contained in molecules excited by the absorption of light should not be available for oxidation-reduction” (p. 152). In this model, the reducing compounds are then used to fix and reduce , and the energy for these reactions comes from the large amounts of free energy released by the reduction reactions.
How is this information explained by our modern understanding of photosynthesis?
Chlorophyll contains a ion, which is known to be an essential cofactor for many enzymes that catalyze phosphorylation and dephosphorylation reactions.
A crude “chlorophyll protein” isolated from photo-synthetic cells showed phosphorylating activity.
The phosphorylating activity of the “chlorophyll protein” was inhibited by light.
The levels of several different phosphorylated compounds in photosynthetic cells changed dramatically in response to light exposure. (Emerson and coworkers were not able to identify the specific compounds involved.)
As it turned out, the Emerson and Rabinowitch models were both partly correct and partly incorrect.
Explain how the two models relate to our current model of photosynthesis.
In his rejection of the Emerson model, Rabinowitch went on to say: “The difficulty of the phosphate storage theory appears most clearly when one considers the fact that, in weak light, eight or ten quanta of light are sufficient to reduce one molecule of carbon dioxide. If each quantum should produce one molecule of high-energy phosphate, the accumulated energy would be only 80–100 kcal per Einstein — while photosynthesis requires at least 112 kcal per mole, and probably more, because of losses in irreversible partial reactions” (p. 228).
How does Rabinowitch’s value of 8 to 10 photons per molecule of reduced compare with the value accepted today?
How would you rebut Rabinowitch’s argument, based on our current knowledge about photosynthesis?
References
- Emerson, R.L., J.F. Stauffer, and W.W. Umbreit. 1944. Relationships between phosphorylation and photosynthesis in Chlorella. Am. J. Botany 31:107–120.
- Rabinowitch, E.I. 1945. Photosynthesis and Related Processes, Vol. I. New York: Interscience Publishers.