The discovery in 1948, by Eugene Kennedy and Albert Lehninger, that mitochondria are the site of oxidative phosphorylation in eukaryotes marked the beginning of the enzymological studies of biological energy transductions. Mitochondria, like gram-negative bacteria, have two membranes (Fig. 19-2a). The outer mitochondrial membrane is readily permeable to small molecules and ions, which move freely through transmembrane channels formed by a family of integral membrane proteins called porins. The inner membrane is impermeable to most small molecules and ions, including protons ; the only species that cross this membrane do so through specific transporters. The inner membrane bears the components of the respiratory chain and ATP synthase.
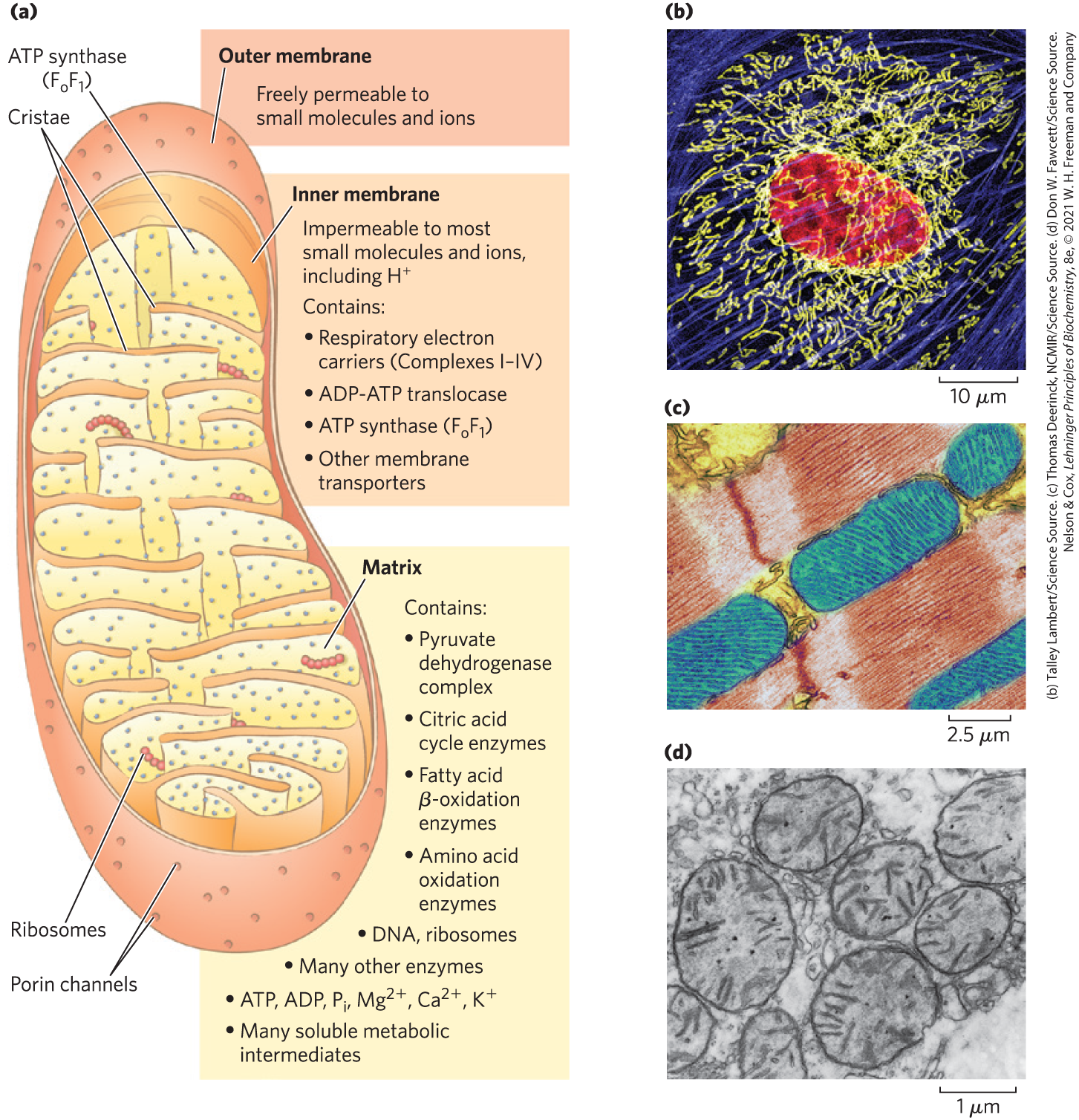
FIGURE 19-2 Biochemical anatomy of a mitochondrion. (a) The outer membrane has pores that make it permeable to small molecules and ions, but not to proteins. The cristae provide a very large surface area. The inner membrane of a single liver mitochondrion may have more than 10,000 sets of electron-transfer systems (respiratory chains) and ATP synthase molecules, distributed over the membrane surface. (b) A typical animal cell has hundreds or thousands of mitochondria. This endothelial cell from bovine pulmonary artery was stained with fluorescent probes for actin (blue), for DNA (red), and for mitochondria (yellow). Notice the variability in length of the mitochondria. (c) The mitochondria of heart muscle (blue in this colorized electron micrograph) have more profuse cristae and thus a much larger area of inner membrane, with more than three times as many sets of respiratory chains as (d) liver mitochondria. Muscle and liver mitochondria are about the size of a bacterium — long. The mitochondria of invertebrates, plants, and microbial eukaryotes are similar to those shown here, but with much variation in size, shape, and degree of convolution of the inner membrane.
The mitochondrial matrix, enclosed by the inner membrane, contains the pyruvate dehydrogenase complex and the enzymes of the citric acid cycle, the fatty acid β-oxidation pathway, and the pathways of amino acid oxidation — all the pathways of fuel oxidation except glycolysis, which takes place in the cytosol. The selectively permeable inner mitochondrial membrane segregates the intermediates and enzymes of cytosolic metabolic pathways from those of metabolic processes occurring in the matrix. However, specific transporters carry pyruvate, fatty acids, and amino acids or their α-keto derivatives into the matrix for access to the machinery of the citric acid cycle. ADP and are specifically transported into the matrix as newly synthesized ATP is transported out. Mammalian mitochondria have about 1,200 proteins, according to current best estimates. The functions of up to 25% of these remain partly or entirely enigmatic.
The bean-shaped representation of a mitochondrion in Figure 19-2a is an oversimplification, derived in part from early studies in which thin sections of cells were observed in the electron microscope. Three-dimensional images obtained either by reconstruction from serial sections or by confocal microscopy reveal great variation in mitochondrial size and shape. In living cells stained with mitochondrion-specific fluorescent dyes, large numbers of variously shaped mitochondria are seen, clustered about the nucleus (Fig. 19-2b).
Tissues with a high demand for aerobic metabolism (brain, skeletal and heart muscle, liver, and eye, for example) contain many hundreds or thousands of mitochondria per cell, and in general, mitochondria of cells with high metabolic activity have more, and more densely packed, convolutions, or cristae (Fig. 19-2c, d). During cell growth and division, mitochondria, like bacteria, divide by fission, and under some circumstances individual mitochondria fuse to form larger, more-extended structures. Stressful conditions, such as the presence of electron-transfer inhibitors or mutations in an electron carrier, trigger mitochondrial fission and sometimes mitophagy — the breakdown of mitochondria and recycling of the amino acids, nucleotides, and lipids released. As stress is relieved, small mitochondria fuse to form long, thin, tubular organelles.
Electrons Are Funneled to Universal Electron Acceptors
Oxidative phosphorylation begins with the entry of electrons into the series of electron carriers called the respiratory chain. Most of these electrons arise from the action of dehydrogenases that collect electrons from catabolic pathways and funnel them into universal electron acceptors — nicotinamide nucleotides ( or ) or flavin nucleotides (FMN or FAD).
Nicotinamide nucleotide–linked dehydrogenases catalyze reversible reactions of the following general types:
Most dehydrogenases that act in catabolism are specific for as electron acceptor (Table 19-1). Some are in the cytosol, many are in mitochondria, and still others have mitochondrial and cytosolic isozymes.
TABLE 19-1 Some Important Reactions Catalyzed by -Linked Dehydrogenases
-linked dehydrogenases remove two hydrogen atoms from their substrates. One of these is transferred as a hydride ion to , and the other is released as in the medium (see Fig. 13-24). NADH and NADPH are water-soluble electron carriers that associate reversibly with dehydrogenases. About 70% of the cellular NAD pool is in the mitochondria. NADH carries electrons from catabolic reactions to their point of entry into the respiratory chain, the NADH dehydrogenase complex described below. NADPH is primarily involved in biosynthetic (anabolic) reactions, and much of it is concentrated in the cytosol. Cells maintain separate pools of NADPH and NADH, with different redox potentials. This is accomplished by holding the ratio [reduced form]/[oxidized form] relatively high for NADPH and relatively low for NADH. Neither NADH nor NADPH can cross the inner mitochondrial membrane, but the electrons they carry can be shuttled across indirectly, as we shall see.
Flavoproteins contain a very tightly, sometimes covalently, bound flavin nucleotide, either FMN or FAD (see Fig. 13-27). The oxidized flavin nucleotide can accept either one electron (yielding the semiquinone form) or two (yielding or ). Electron transfer occurs because the flavoprotein has a higher reduction potential than the compound oxidized. Recall that reduction potential is a quantitative measure of the relative tendency of a given chemical species to accept electrons in an oxidation-reduction reaction (p. 490). The standard reduction potential of a flavin nucleotide, unlike that of NAD or NADP, depends on the protein with which it is associated. Local interactions with functional groups in the protein distort the electron orbitals in the flavin ring, changing the relative stabilities of oxidized and reduced forms. The relevant standard reduction potential is therefore that of the particular flavoprotein, not that of isolated FAD or FMN. The flavin nucleotide should be considered part of the flavoprotein’s active site rather than a reactant or product in the electron-transfer reaction. Because flavoproteins can participate in either one- or two-electron transfers, they can serve as intermediates between reactions in which two electrons are donated (as in dehydrogenations) and those in which only one electron is accepted (as in the reduction of a quinone to a hydroquinone, described below).
Electrons Pass through a Series of Membrane-Bound Carriers
The mitochondrial respiratory chain consists of a series of sequentially acting electron carriers, most of which are integral proteins with prosthetic groups capable of accepting and donating either one or two electrons. Three types of electron transfers occur in oxidative phosphorylation: (1) direct transfer of electrons, as in the reduction of to ; (2) transfer as a hydrogen atom ; and (3) transfer as a hydride ion , which bears two electrons. The term reducing equivalent is used to designate a single electron equivalent transferred in an oxidation-reduction reaction.
In addition to NAD and flavoproteins, three other types of electron-carrying molecules function in the respiratory chain: a hydrophobic quinone called ubiquinone, and two different types of iron-containing proteins, cytochromes and iron-sulfur proteins. Ubiquinone (also called coenzyme Q, or simply Q) is a lipid-soluble benzoquinone with a long isoprenoid side chain (Fig. 19-3). Ubiquinone can accept one electron to become the semiquinone radical ( or ubisemiquinone) or two electrons to form ubiquinol and, like flavoprotein carriers, it can act at the junction between a two-electron donor and a one-electron acceptor. Small and hydrophobic, ubiquinone is not bound to proteins but instead is freely diffusible within the lipid bilayer of the inner mitochondrial membrane and can shuttle reducing equivalents between other, less mobile electron carriers in the membrane. And because it carries both electrons and protons, it plays a central role in coupling electron flow to proton movement.
FIGURE 19-3 Ubiquinone (Q, or coenzyme Q). Complete reduction of ubiquinone requires two electrons and two protons, and occurs in two steps through the semiquinone radical intermediate.
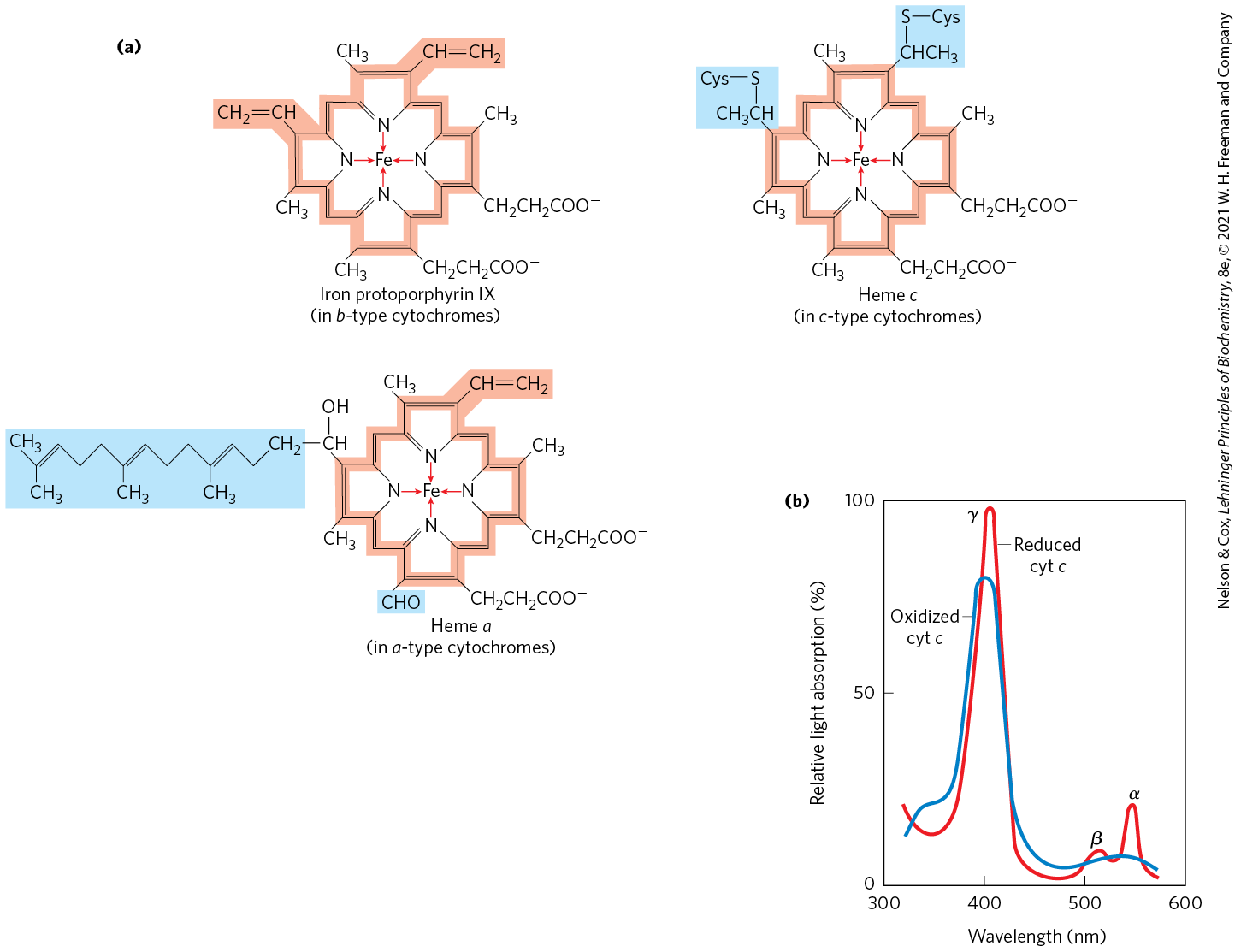
The cytochromes are proteins with characteristic strong absorption of visible light, due to their iron-containing heme prosthetic groups (Fig. 19-4a). Mitochondria contain three classes of cytochromes, designated a, b, and c, which are distinguished by differences in their light-absorption spectra. Each type of cytochrome in its reduced state has three absorption bands in the visible range (Fig. 19-4b). The longest-wavelength band is near 600 nm in type a cytochromes, near 560 nm in type b, and near 550 nm in type c. To distinguish among closely related cytochromes of one type, the exact absorption maximum is sometimes used in the names, as in cytochrome .
FIGURE 19-4 Prosthetic groups of cytochromes. (a) Each group consists of four five-membered, nitrogen-containing rings in a cyclic structure called a porphyrin. The four nitrogen atoms are coordinated with a central Fe ion, either or . Iron protoporphyrin IX is found in b-type cytochromes and in hemoglobin and myoglobin (see Fig. 5-1b). Heme c is covalently bound to the protein of cytochrome c through thioether bonds to two Cys residues. Heme a, found in a-type cytochromes, has a long isoprenoid tail attached to one of the five-membered rings. The conjugated double-bond system (shaded light red) of the porphyrin ring has delocalized π electrons that are relatively easily excited by photons with the wavelengths of visible light, which accounts for the strong absorption by hemes (and related compounds) in the visible region of the spectrum. (b) Absorption spectra of cytochrome c (cyt c) in its oxidized (blue) and reduced (red) forms. The characteristic α, β, and γ bands of the reduced form are labeled.
The hemes of a and b cytochromes are tightly, but not covalently, bound to their associated proteins; the hemes of c-type cytochromes are covalently attached through Cys residues (Fig. 19-4). As with the flavoproteins, the standard reduction potential of the heme iron atom of a cytochrome depends on its interaction with protein side chains and is therefore different for each cytochrome. The cytochromes of type a and b and some of type c are integral proteins of the inner mitochondrial membrane. One striking exception is the soluble cytochrome c that associates through electrostatic interactions with the outer surface of the inner membrane.
In iron-sulfur proteins, the iron is present not in heme but in association with inorganic sulfur atoms or with the sulfur atoms of Cys residues in the protein, or both. These iron-sulfur (Fe-S) centers range from simple structures with a single Fe atom coordinated to four Cys OSH groups to more complex Fe-S centers with two or four Fe atoms (Fig. 19-5). Rieske iron-sulfur proteins (named after their discoverer, John S. Rieske) are a variation on this theme, in which one Fe atom is coordinated to two His residues rather than two Cys residues. All iron-sulfur proteins participate in one-electron transfers in which one iron atom of the Fe-S cluster is oxidized or reduced. At least eight Fe-S proteins function in mitochondrial electron transfer. The reduction potential of Fe-S proteins varies from to , depending on the microenvironment of the iron within the protein.
FIGURE 19-5 Iron-sulfur centers. The Fe-S centers of iron-sulfur proteins may be as simple as shown in (a), with a single Fe ion surrounded by the S atoms of four Cys residues; inorganic S is yellow and the S of Cys is orange. Other centers include both inorganic and Cys S atoms, as in (b) 2Fe-2S or (c) 4Fe-4S centers. (d) The ferredoxin of the cyanobacterium Anabaena 7120 has one 2Fe-2S center. (Note that in these designations, only the inorganic S atoms are counted. For example, in the 2Fe-2S center (b), each Fe ion is actually surrounded by four S atoms.) The exact standard reduction potential of the iron in these centers depends on the type of center and its interaction with the associated protein. [(d) Data from PDB ID 1FRD, B. L. Jacobson et al., Biochemistry 32:6788, 1993.]
In the overall reaction catalyzed by the mitochondrial respiratory chain, electrons move from NADH, succinate, or some other primary electron donor through flavoproteins, ubiquinone, iron-sulfur proteins, and cytochromes, and finally to . A look at the methods used to determine the sequence in which the carriers act is instructive, as the same general approaches have been used to study other electron-transfer chains, such as those of chloroplasts (see Fig. 20-12).
First, the standard reduction potentials of the individual electron carriers have been determined experimentally (Table 19-2). Electrons tend to flow spontaneously from carriers of lower to carriers of higher . The order of carriers deduced by this method is cytochrome . Note, however, that the order of standard reduction potentials is not necessarily the same as the order of actual reduction potentials under cellular conditions, which depend on the concentrations of reduced and oxidized forms (see Eqn 13-5, p. 491). A second method for determining the sequence of electron carriers involves reducing the entire chain of carriers experimentally by providing an electron source but no electron acceptor (no ). When is suddenly introduced into the system, the rate at which each electron carrier becomes oxidized, measured spectroscopically, reveals the order in which the carriers function. The carrier nearest (at the end of the chain) gives up its electrons first, the second carrier from the end is oxidized next, and so on. Such experiments have confirmed the sequence deduced from standard reduction potentials.
TABLE 19-2 Standard Reduction Potentials of Respiratory Chain and Related Electron Carriers
Redox reaction (half-reaction)
(V)
−0.414
−0.320
−0.324
−0.30
0.045
0.077
0.22
0.254
0.29
0.35
0.817
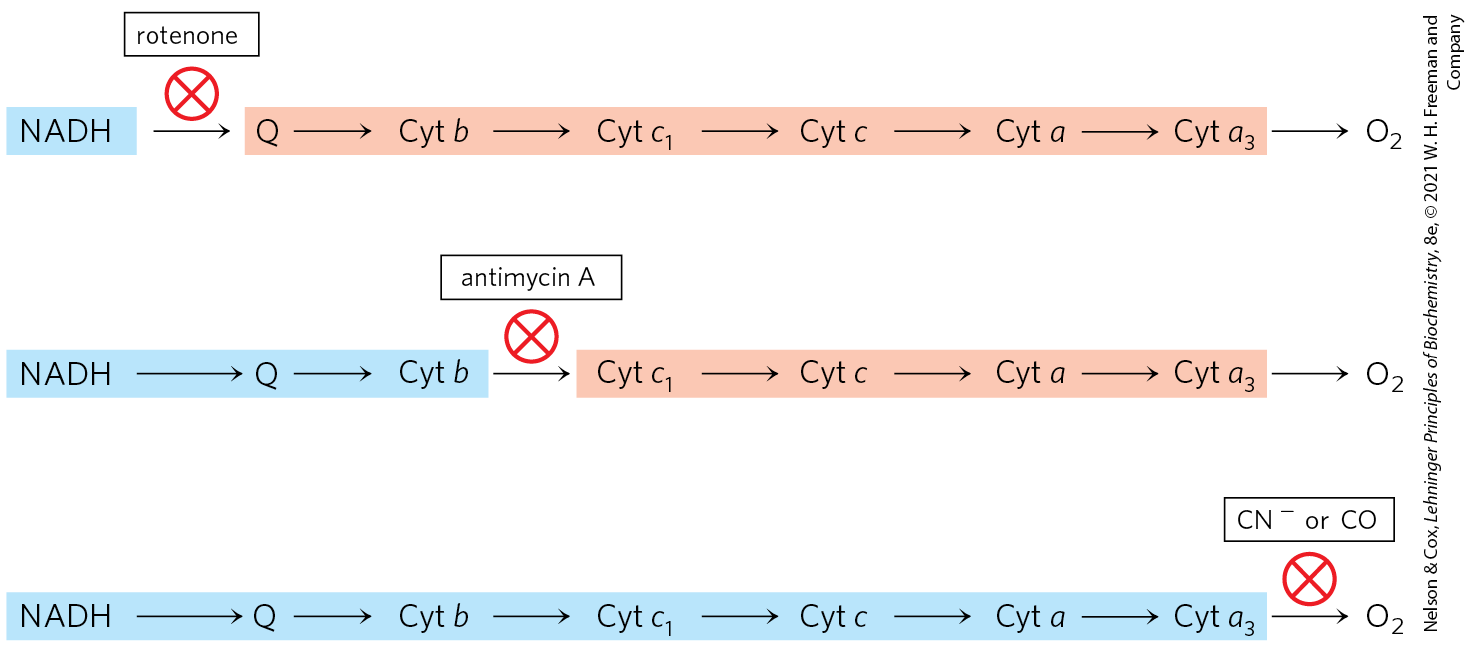
In a final confirmation, agents that inhibit the flow of electrons through the chain have been used in combination with measurements of the degree of oxidation of each carrier. In the presence of and an electron donor, carriers that function before the inhibited step become fully reduced, and those that function after this step are completely oxidized (Fig. 19-6). By using several inhibitors that block different steps in the chain, investigators have determined the entire sequence; it is the same as deduced in the first two approaches.
FIGURE 19-6 Method for determining the sequence of electron carriers. This method measures the effects of inhibitors of electron transfer on the oxidation state of each carrier. In the presence of an electron donor and , each inhibitor causes a characteristic pattern of oxidized/reduced carriers: those before the block become reduced (blue), and those after the block become oxidized (light red).
Electron Carriers Function in Multienzyme Complexes
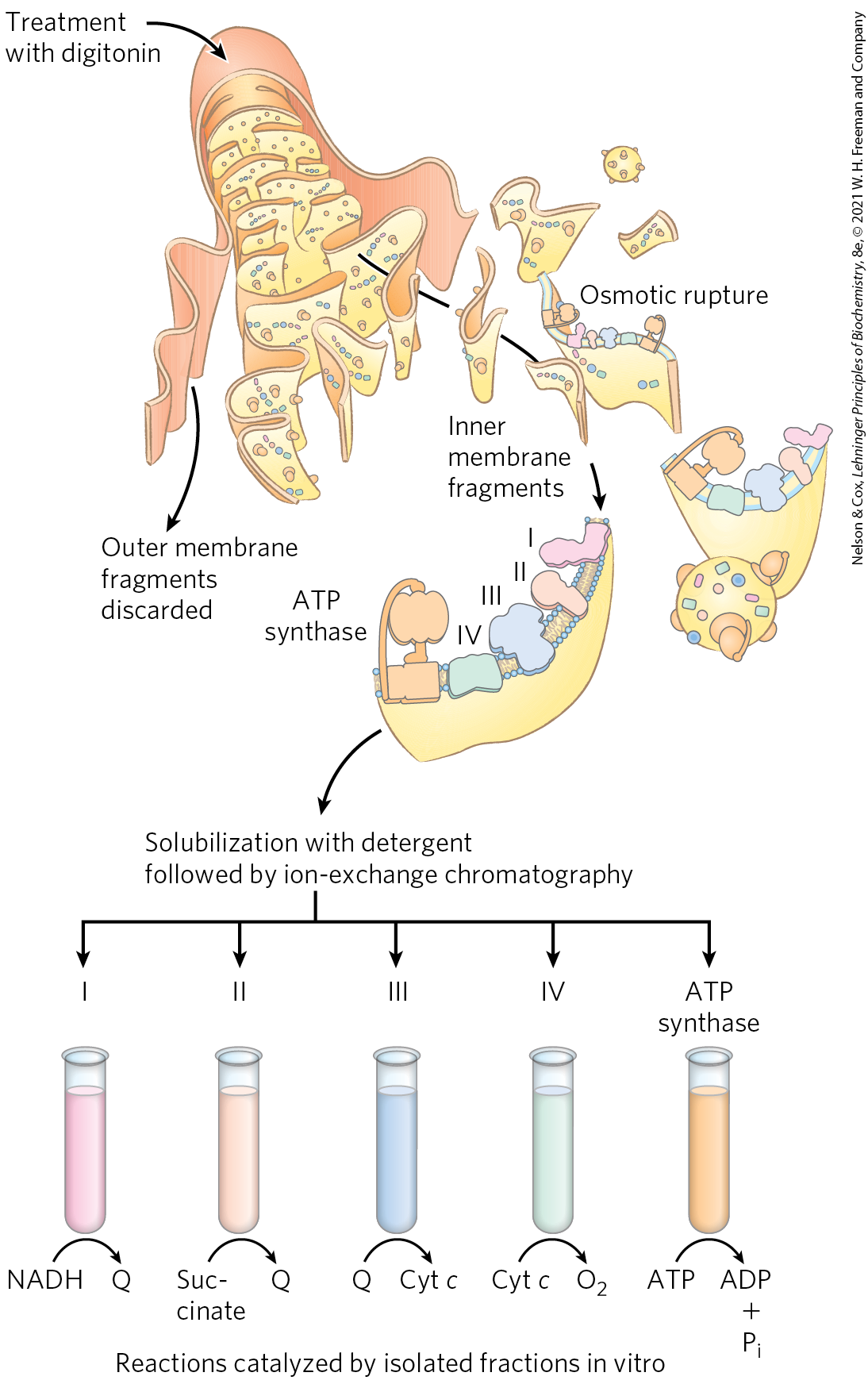
The electron carriers of the respiratory chain are organized into membrane-embedded supramolecular complexes that can be physically separated. Gentle treatment of the inner mitochondrial membrane with detergents allows the resolution of four unique electron-carrier complexes, each capable of catalyzing electron transfer through a portion of the chain (Fig. 19-7; Table 19-3). Complexes I and II catalyze electron transfer to ubiquinone from two different electron donors: NADH (Complex I) and succinate (Complex II). Complex III carries electrons from reduced ubiquinone to cytochrome c, and Complex IV completes the sequence by transferring electrons from cytochrome c to .
FIGURE 19-7 Separation of functional complexes of the respiratory chain. The outer mitochondrial membrane is first removed by treatment with the detergent digitonin. Fragments of inner membrane are then obtained by osmotic rupture of the membrane, and the fragments are gently dissolved in a second detergent. The resulting mixture of inner membrane proteins is resolved by ion-exchange chromatography into several complexes (I through IV) of the respiratory chain, each with its unique protein composition (see Table 19-3), and the enzyme ATP synthase (sometimes called Complex V). The isolated Complexes I through IV catalyze electron transfers between donors (NADH and succinate), intermediate carriers (Q and cytochrome c), and , as shown. In vitro, isolated ATP synthase has only ATP-hydrolyzing (ATPase), not ATP-synthesizing, activity.
TABLE 19-3 The Protein Components of the Mitochondrial Respiratory Chain
aNumber of subunits in bacterial complexes is shown in parentheses.
bMass and subunit data are for the monomeric form.
cCytochrome c is not part of an enzyme complex; it moves between Complexes III and IV as a freely soluble protein.
We now look in more detail at the structure and function of each complex of the mitochondrial respiratory chain.
Complex I: NADH to Ubiquinone
In mammals, Complex I, also called NADH:ubiquinone oxidoreductase or NADH dehydrogenase, is a large enzyme composed of 45 different polypeptide chains, including an FMN-containing flavoprotein and at least 8 iron-sulfur centers. Complex I is L-shaped, with one arm embedded in the inner membrane and the other extending into the matrix. Comparative studies of Complex I in bacteria and other organisms show that 7 polypeptides in the membrane arm and 7 in the matrix arm are conserved and essential (Fig. 19-8).
FIGURE 19-8 Structure of Complex I (NADH:ubiquinone oxidoreductase). Complex I catalyzes the transfer of a hydride ion from NADH to FMN. From the FMN, two electrons pass through a series of Fe-S centers to the Fe-S center N-2 in the matrix arm of the complex. Electron transfer from N-2 to ubiquinone on the membrane arm forms , which diffuses into the lipid bilayer. The protons travel a path dictated by subunit conformation changes triggered by the electron flow. Proton flux produces an electrochemical potential across the inner mitochondrial membrane (n side negative, p side positive). Three of the membrane subunits (subunits Nqo12, Nqo13, and Nqo14) are structurally related to a known antiporter, and the path of proton movement may be similar in both cases. The fourth putative proton pathway is through an integral subunit closest to the Q-binding site. A long helix (not visible in this view) lying along the surface of the membrane arm may coordinate the action of all four proton pumps when Q is reduced. [Data from PDB ID 4HEA, R. Baradaran et al., Nature 494:443, 2013.]
Complex I catalyzes two simultaneous and obligately coupled processes: (1) the exergonic transfer to ubiquinone of a hydride ion from NADH and a proton from the matrix, expressed by
(19-1)
and (2) the endergonic transfer of four protons from the matrix to the intermembrane space. Protons are moved against a transmembrane proton gradient in this process. Complex I is therefore a proton pump driven by the energy of electron transfer, and the reaction it catalyzes is vectorial: it moves protons in a specific direction from one location (the matrix, which becomes negatively charged with the departure of protons) to another (the intermembrane space, which becomes positively charged). To emphasize the vectorial nature of the process, the overall reaction is often written with subscripts that indicate the location of the protons: p for the positive side of the inner membrane (the intermembrane space), n for the negative side (the matrix):
(19-2)
Amytal (a barbiturate drug), rotenone (a plant product commonly used as an insecticide), and piericidin A (an antibiotic) inhibit electron flow from the Fe-S centers of Complex I to ubiquinone (Table 19-4) and therefore block the overall process of oxidative phosphorylation.
TABLE 19-4 Agents That Interfere with Oxidative Phosphorylation
Three of the seven integral protein subunits of the membrane arm are related to a antiporter and are believed to be responsible for pumping three protons; a fourth subunit in the membrane arm, that nearest the Q-binding site, is probably responsible for pumping the fourth proton (Fig. 19-8).
How is the reduction of ubiquinone coupled to proton pumping? Reduction of Q occurs far away from the membrane arm of the protein, where proton pumping occurs, so the coupling is clearly indirect. The high-resolution view of Complex I from crystallographic and cryo-EM studies suggests that reduction of Q is coupled to a long-range conformational change conducted to all subunits along the hydrophilic core of the transmembrane arm. It seems likely that all four protons are pumped simultaneously, so that the energy from a strongly exergonic reaction (Q reduction) is broken into smaller packets, a common strategy employed by living organisms.
Complex II: Succinate to Ubiquinone
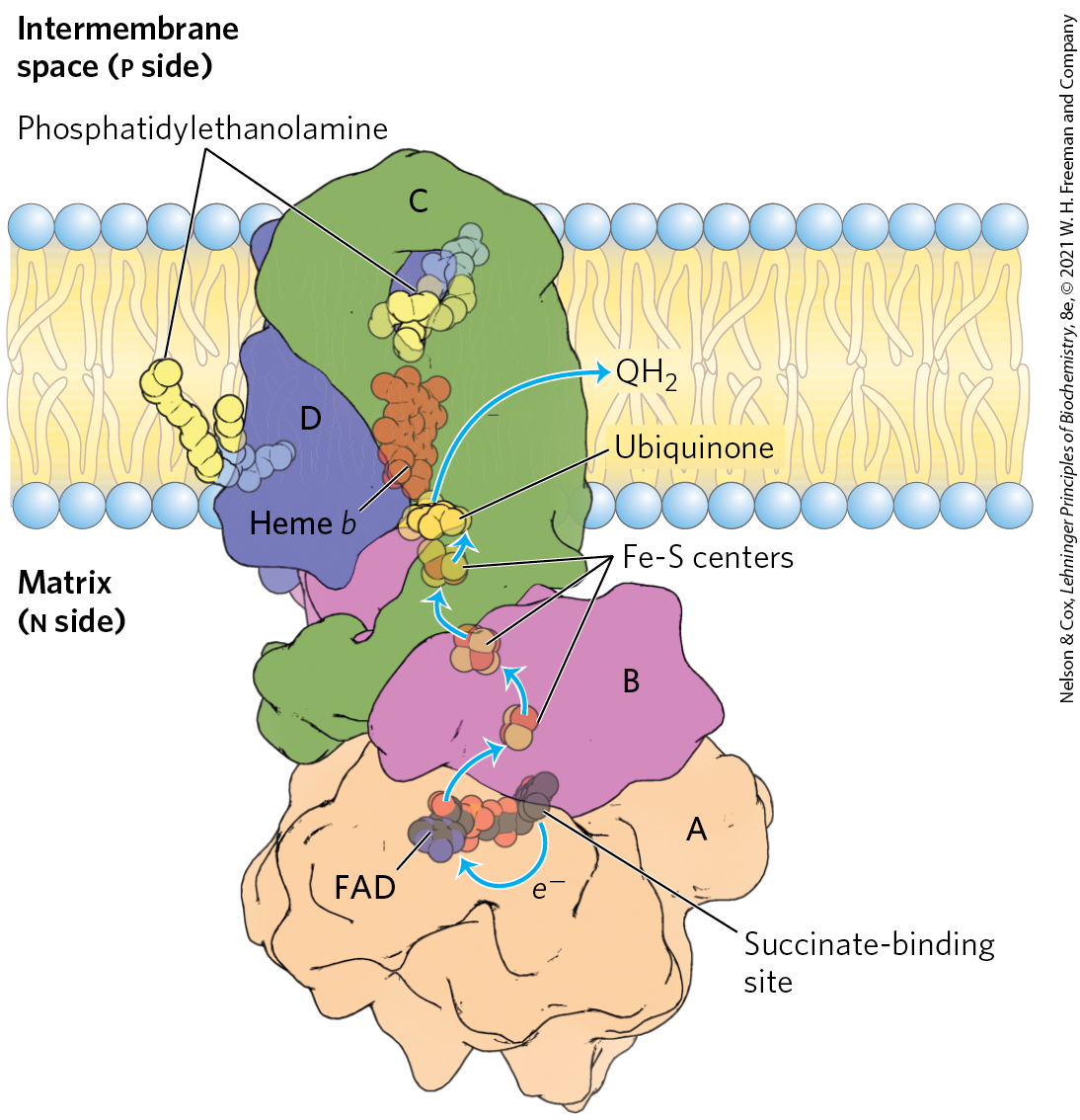
We encountered Complex II in Chapter 16 as succinate dehydrogenase, the only membrane-bound enzyme in the citric acid cycle (p. 586). Complex II couples the oxidation of succinate at one site with the reduction of ubiquinone at another site about 40 Å away. Although smaller and simpler than Complex I, Complex II contains five prosthetic groups of two types and four different protein subunits (Fig. 19-9). Subunits C and D are integral membrane proteins, each with three transmembrane helices. They contain a heme group, heme b, and a binding site for Q, the final electron acceptor in the reaction catalyzed by Complex II. Subunits A and B extend into the matrix; they contain three 2Fe-2S centers, bound FAD, and a binding site for the substrate, succinate. Although the overall path of electron transfer is long (from the succinate-binding site to FAD, then through the Fe-S centers to the Q-binding site), none of the individual electron-transfer distances exceeds about 11 Å — a reasonable distance for rapid electron transfer (Fig. 19-9). Electron transfer through Complex II is not accompanied by proton pumping across the inner membrane, although the produced by succinate oxidation will be used by Complex III to drive proton transfer. Because Complex II functions in the citric acid cycle, factors that affect its activity (such as the availability of oxidized Q) probably serve to coordinate that cycle with mitochondrial electron transfer.
FIGURE 19-9 Structure of Complex II (succinate dehydrogenase). This complex (porcine) has two transmembrane subunits, C and D; subunits A and B extend into the matrix. Just behind the FAD in subunit A is the binding site for succinate. Subunit B has three Fe-S centers, ubiquinone is bound to subunit B, and heme b is sandwiched between subunits C and D. Two phosphatidylethanolamine molecules are so tightly bound to subunit D that they show up in the crystal structure. Electrons move (blue arrows) from succinate to FAD, then through the three Fe-S centers to ubiquinone. The heme b is not on the main path of electron transfer but protects against the formation of reactive oxygen species (ROS) by electrons that go astray. [Data from PDB ID 1ZOY, F. Sun et al., Cell 121:1043, 2005.]
The heme b of Complex II is apparently not in the direct path of electron transfer; it serves instead to reduce the frequency with which electrons “leak” out of the system, moving from succinate to molecular oxygen to produce the reactive oxygen species (ROS) hydrogen peroxide and the superoxide radical , as described below. Some individuals with point mutations in Complex II subunits near heme b or the ubiquinone-binding site suffer from hereditary paraganglioma, characterized by benign tumors of the head and neck, commonly in the carotid body, an organ that senses levels in the blood. These mutations result in greater production of ROS, which cause DNA damage and genome instability that can lead to cancer. Mutations that affect the succinate-binding region in Complex II may lead to degenerative changes in the central nervous system, and some mutations are associated with tumors of the adrenal medulla.
Complex III: Ubiquinone to Cytochrome c
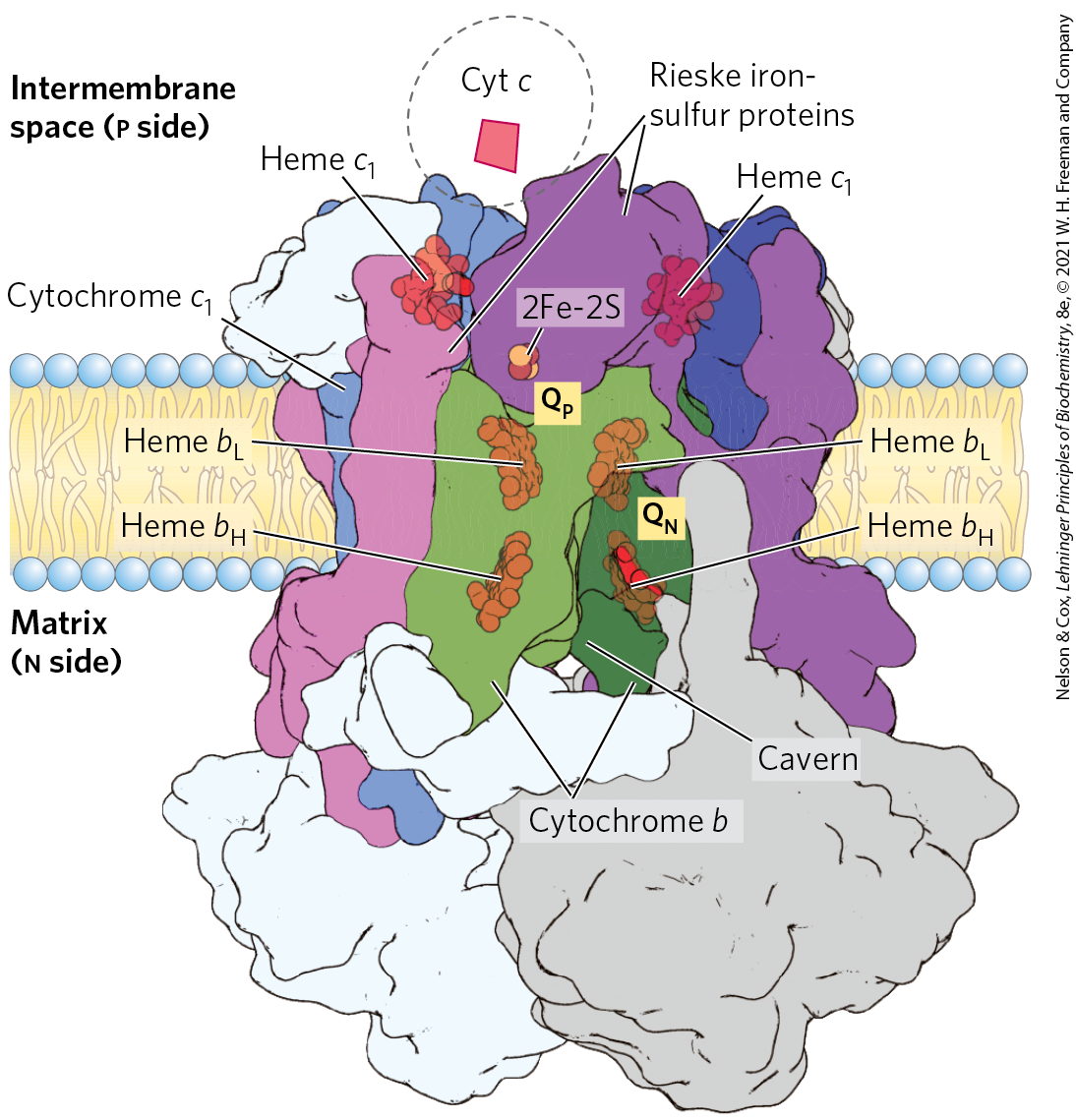
Electrons from reduced ubiquinone (ubiquinol, ) pass through two more large protein complexes in the inner mitochondrial membrane before reaching the ultimate electron acceptor, . Complex III (also called cytochrome complex or ubiquinone:cytochrome c oxidoreductase) couples the transfer of electrons from ubiquinol to cytochrome c with the vectorial transport of protons from the matrix to the intermembrane space. The functional unit of Complex III (Fig. 19-10) is a dimer. Each monomer consists of three proteins central to the action of the complex: cytochrome b, cytochrome , and the Rieske iron-sulfur protein. (Several other proteins associated with Complex III in vertebrates are not conserved across the phyla and presumably play subsidiary roles.) The two cytochrome b monomers surround a cavern in the middle of the membrane, in which ubiquinone is free to move from the matrix side of the membrane (site on one monomer) to the intermembrane space (site on the other monomer) as it shuttles electrons and protons across the inner mitochondrial membrane.
FIGURE 19-10 Structure of Complex III (cytochrome complex). The complex (bovine) is a dimer of identical monomers, each with 11 different subunits. The functional core of each monomer consists of three subunits: cytochrome b (green), with its two hemes ; the Rieske iron-sulfur protein (purple), with its 2Fe-2S centers; and cytochrome (blue), with its heme. Cytochrome and the Rieske iron-sulfur protein project from the p surface and can interact with cytochrome c (not part of the functional complex) in the intermembrane space. The complex has two distinct binding sites for ubiquinone, and , which correspond to the sites of inhibition by two drugs that block oxidative phosphorylation. Antimycin A, which blocks electron flow from cytochrome b to cytochrome , specifically from heme to Q, binds at , close to heme on the n (matrix) side of the membrane. Myxothiazol, which prevents electron flow from to the Rieske iron-sulfur protein, binds at , near the 2Fe-2S center and heme on the p side. The dimeric structure is essential to the function of Complex III. The interface between monomers forms two caverns, each containing a site from one monomer and a site from the other. The ubiquinone intermediates move within these sheltered caverns. [Data from PDB ID 1BGY, S. Iwata et al., Science 281:64, 1998.]
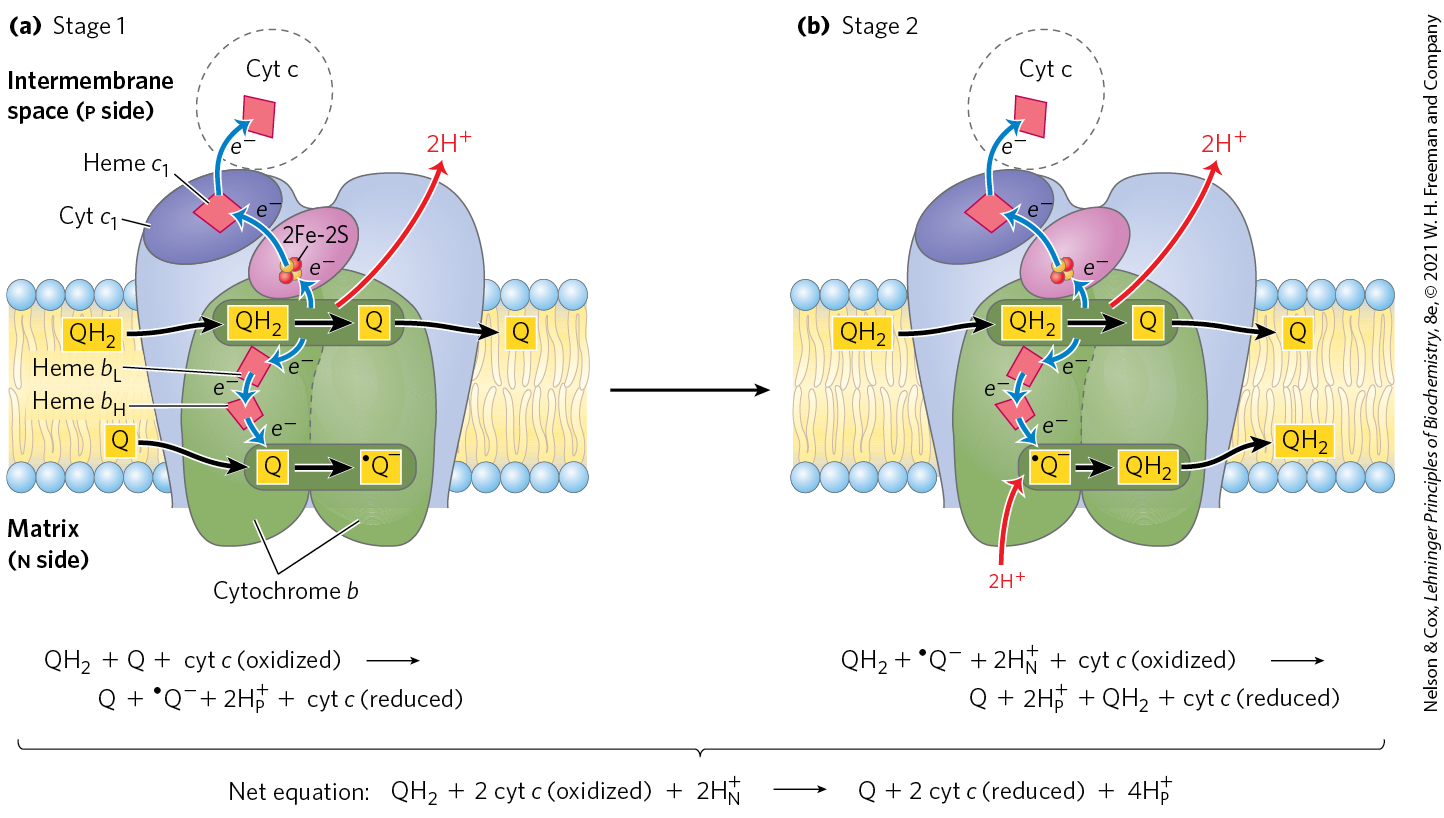
To account for the role of Q in energy conservation, Mitchell proposed the Q cycle (Fig. 19-11). As electrons move from through Complex III, is oxidized with the release of protons on one side of the membrane (at ), while at the other site , Q is reduced and protons are taken up.
FIGURE 19-11 The Q cycle, shown in two stages. The path of electrons through Complex III is shown with blue arrows; the movement of various forms of ubiquinone, with black arrows. (a) In the first stage, Q on the n side is reduced to the semiquinone radical, which moves back into position (dotted line) to accept another electron. (b) In the second stage, the semiquinone radical is converted to . Meanwhile, on the p side of the membrane, two molecules of are oxidized to Q, releasing two protons per Q molecule (four protons in all) into the intermembrane space. Each donates one electron (via the Rieske Fe-S center) to cytochrome , and one electron (via cytochrome b) to a molecule of Q near the n side, reducing it in two steps to . This reduction also consumes two protons per Q, which are taken up from the matrix (n side). Reduced cytochrome passes electrons one at a time to cytochrome c, which dissociates and carries electrons to Complex IV. In each cycle, one reduction of Q at the site is coupled with two oxidations of at the site by consuming two protons from the matrix and releasing four protons into the intermembrane space.
The Q cycle is most easily understood as occurring in two stages, with two active sites where ubiquinone is either oxidized or reduced. In both stages, one is oxidized at active site 1, shedding two and two electrons. The protons are released into the intermembrane space. The two electrons take different paths, with one reducing cytochrome c and the other reducing a molecule of Q at active site 2. Two electrons are required at active site 2 to fully reduce the Q to , one in each stage. Reducing one Q at one site while oxidizing two at another may seem counterproductive at first glance. However, the two processes have complementary functions. The oxidation of two is moving four protons to the intermembrane space and two electrons to cytochrome c. At the same time, the reduction of Q at the other site (using the other two electrons from the oxidation of at site 1) is pulling protons from the matrix, creating a net movement of protons from the matrix to the intermembrane space. The produced at active site 2 becomes a substrate for oxidation at active site 1 in subsequent turns of the cycle, and vice versa. The net equation for the redox reactions of the Q cycle is
(19-3)
The Q cycle accommodates the switch between the two-electron carrier ubiquinol (the reduced form of ubiquinone) and the one-electron carriers — hemes and of cytochrome b, and cytochromes and c — and results in the uptake of two protons on the n side and the release of four protons on the p side, per pair of electrons passing through Complex III to cytochrome c. Two of the protons released on the p side are electrogenic; the other two are electroneutral, balanced by the two charges (electrons) passed to cytochrome c on the p side. Although the path of electrons through this segment of the respiratory chain is complicated, the net effect of the transfer is simple: is oxidized to Q, two molecules of cytochrome c are reduced, and two protons are moved from the n side to the p side of the inner mitochondrial membrane.
Cytochrome c is a soluble protein of the intermembrane space, which associates reversibly with the p side of the inner membrane. After its single heme accepts an electron from Complex III, cytochrome c moves in the intermembrane space to Complex IV to donate the electron to a binuclear copper center.
Complex IV: Cytochrome c to
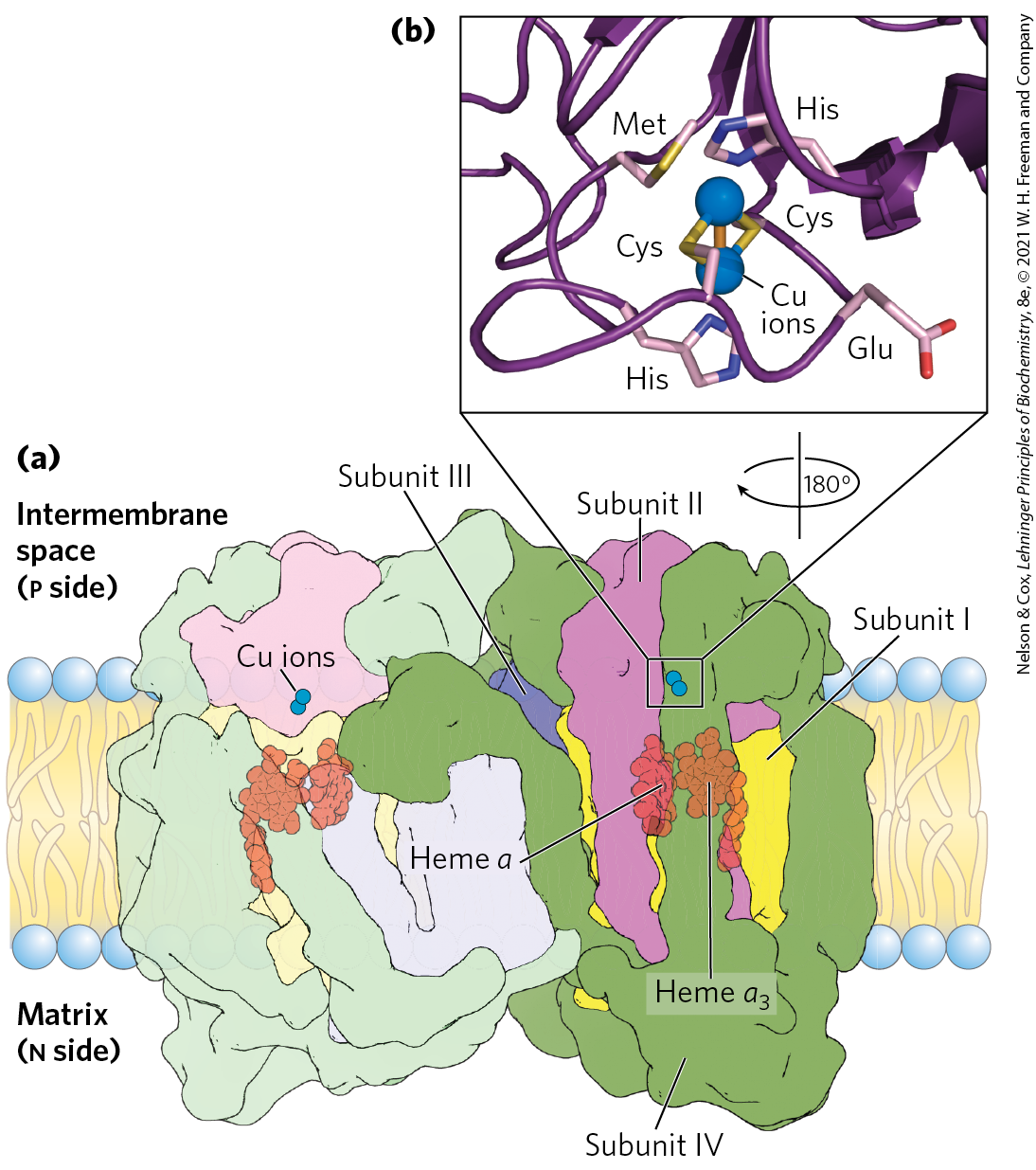
In the final step of the respiratory chain, Complex IV, also called cytochrome oxidase, carries electrons from cytochrome c to molecular oxygen, reducing it to . Complex IV is a large, dimeric enzyme of the inner mitochondrial membrane, each monomer having 13 subunits and of 204,000. Bacteria contain a form that is much simpler, with only 3 or 4 subunits per monomer, but still capable of catalyzing both electron transfer and proton pumping. Comparison of the mitochondrial and bacterial complexes suggests that these 3 subunits have been conserved in evolution; in multicellular organisms, the other 10 subunits contribute to the assembly or stability of Complex IV (Fig. 19-12).
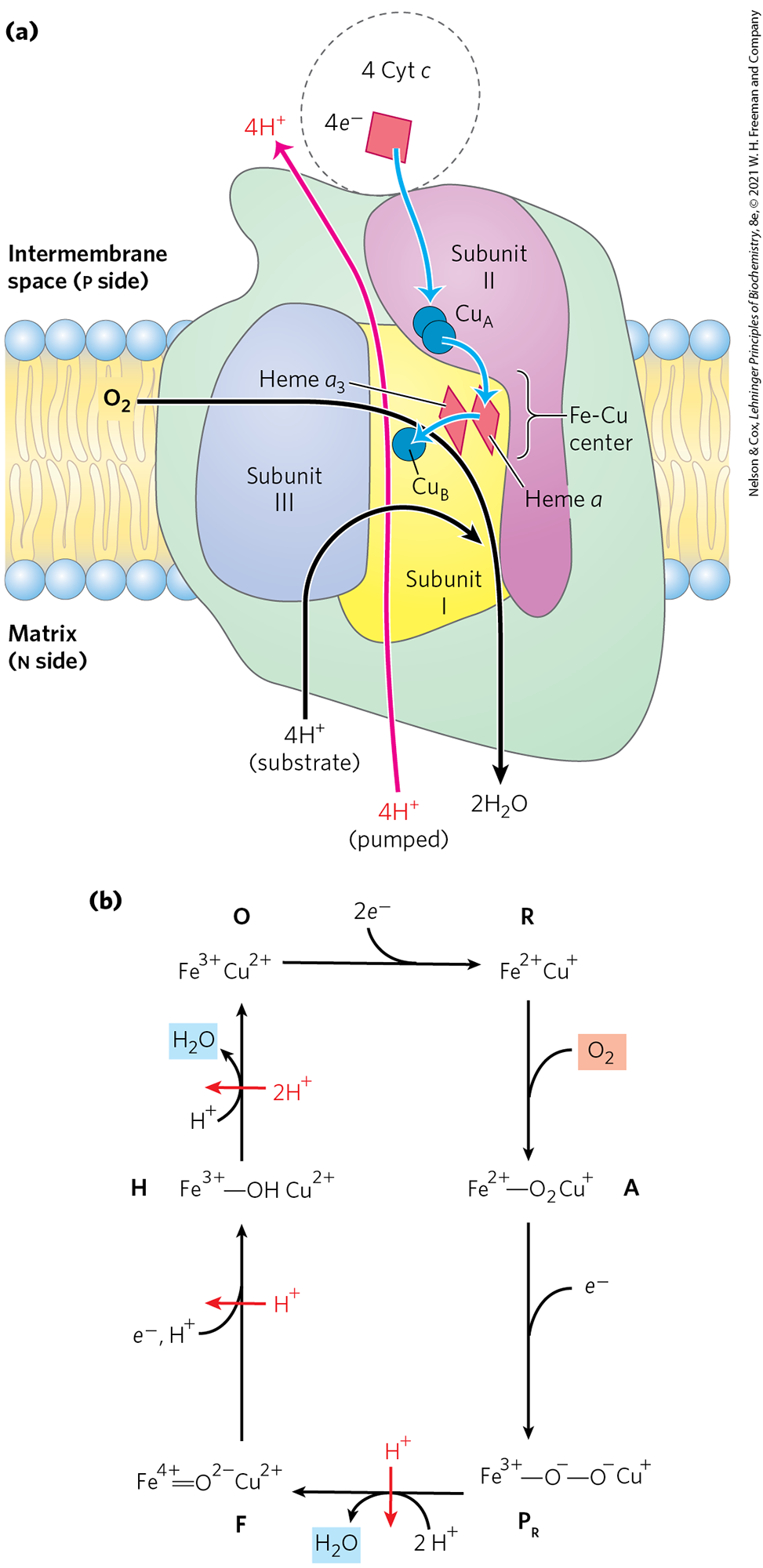
FIGURE 19-12 Structure of Complex IV (cytochrome oxidase). (a) This complex (bovine) has 13 subunits in each identical monomer of its dimeric structure. Subunit I has two heme groups, a and , near a single copper ion, (not visible here). Heme and form a binuclear Fe-Cu center. Subunit II contains two Cu ions complexed with the —SH groups of two Cys residues in a binuclear center, , that resembles the 2Fe-2S centers of iron-sulfur proteins. This binuclear center and the cytochrome c–binding site are located in a domain of subunit II that protrudes from the p side of the inner membrane (into the intermembrane space). Subunit III is essential for rapid proton movement through subunit II. The roles of the other 10 subunits in mammalian Complex IV are not fully understood, although some function in assembly or stabilization of the complex. (b) The binuclear center of . The Cu ions (blue spheres) share electrons equally. When the center is reduced, the ions have the formal charges ; when oxidized, . Six amino acid residues are ligands around the Cu ions: Glu, Met, two His, and two Cys. [Data from PDB ID 1OCC, T. Tsukihara et al., Science 272:1136, 1996.]
Subunit II of Complex IV contains two Cu ions complexed with the —SH groups of two Cys residues in a binuclear center (; Fig. 19-12b) that resembles the 2Fe-2S centers of iron-sulfur proteins. Subunit I contains two heme groups, designated a and , and another copper ion . Heme and form a second binuclear center that accepts electrons from heme a and transfers them to bound to heme . The detailed role of subunit III is not clear, but its presence is essential to Complex IV function.
Electron transfer through Complex IV is from cytochrome c to the center, to heme a, to the heme center, and finally to (Fig. 19-13a). For every four electrons passing through this complex, the enzyme consumes four “substrate” from the matrix (n side) in converting to two . It also uses the energy of this redox reaction to pump four protons outward into the intermembrane space (p side) for each four electrons that pass through, adding to the electrochemical potential produced by redox-driven proton transport through Complexes I and III. The overall reaction catalyzed by Complex IV is
(19-4)
FIGURE 19-13 Path of electrons through Complex IV. (a) For simplicity, only one monomer of the dimeric bovine Complex IV is shown. The three proteins critical to electron flow are subunits I, II, and III. The larger green structure includes the other 10 proteins in each monomer of the dimeric complex. Electron transfer through Complex IV begins with cytochrome c (top). Two molecules of reduced cytochrome c each donate an electron to the binuclear center . From here, electrons pass through heme a to the Fe-Cu center (heme and ). Oxygen now binds to heme and is reduced to its peroxy derivative (; not shown here) by two electrons from the Fe-Cu center. Delivery of two more electrons from cytochrome c (top), for a total of four electrons, converts the to two molecules of water, with consumption of four “substrate” protons from the matrix. At the same time, four protons are pumped from the matrix for every four electrons passing through Complex IV. A simplified reaction sequence is presented in (b). Intermediate complexes O, R, A, , F, and H represent only a prominent subset of the species for which there is experimental evidence, with some steps and intermediate structures still being debated. The four electrons are introduced in separate steps, and is released in two separate steps.
Note that the in this reaction is the final acceptor for electrons originating from the many sources already described, and the stoichiometries between electron sources and molecules consumed help to define the energetics of the systems. In this chapter, stoichiometries are sometimes presented, as here, in terms of one molecule of . For calculation simplicity, the stoichiometries will be presented in terms of in some examples to come.
At Complex IV, is reduced at redox centers that carry only one electron at a time. A reaction scheme is presented in Figure 19-13b. Normally the incompletely reduced oxygen intermediates remain tightly bound to the complex until completely converted to water. However, a small fraction of oxygen intermediates escape. These intermediates are reactive oxygen species that can damage cellular components unless eliminated by defense mechanisms described below.
Mitochondrial Complexes Associate in Respirasomes
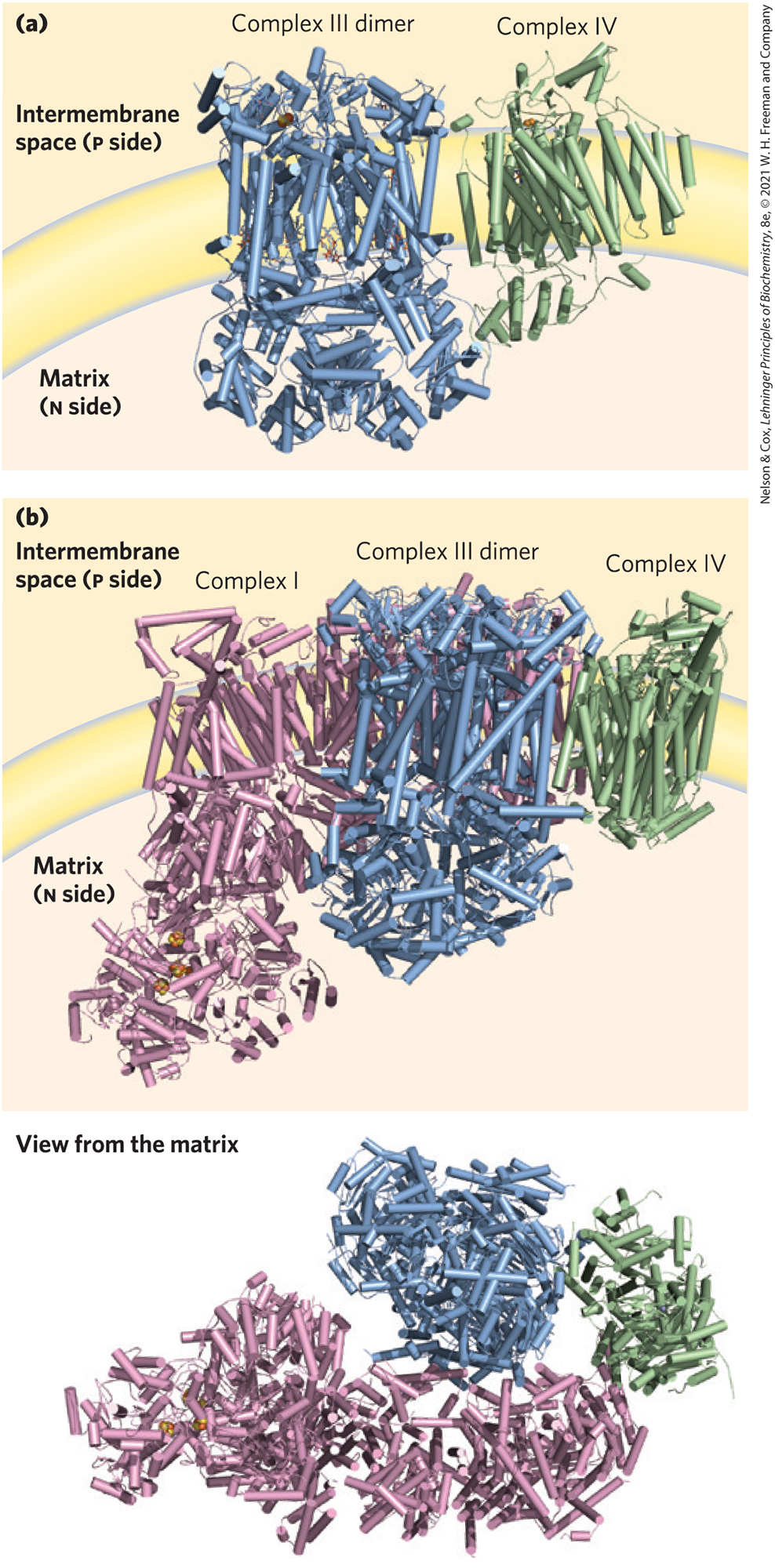
Although the four electron-transferring complexes can be separated in the laboratory, in the intact mitochondrion, three of the four respiratory complexes associate with each other in the inner membrane. Combinations of Complexes I and III, III and IV, and I, III, and IV are formed in organisms ranging from yeast to plants to mammals. The supercomplex containing Complexes I, III, and IV has been called the respirasome. Unlike the other three complexes, Complex II is generally found free-floating within the membrane. Structural characterization of the various supercomplexes has been advanced by cryo-EM (Fig. 19-14). The functional significance of supercomplexes has not been determined. Researchers have suggested that they may facilitate electron transfers or limit the production of reactive oxygen species. Local pools of the electron carriers cytochrome c and ubiquinone are not constrained within supercomplexes, but instead readily diffuse between them.
FIGURE 19-14 A respirasome composed of Complexes I, III, and IV. (a) Purified supercomplexes containing Complexes III and IV (from yeast), as determined by cryo-EM. (b) The structure of a respirasome composed of mammalian (porcine and bovine) Complexes I, III, and IV. Two views are shown. [Data from (a) PDB ID 6GIQ, S. Rathore et al., Nat. Struct. Mol. Biol. 26:50, 2019; (b) PDB ID 5GPN, J. Gu et al., Nature 537:639, 2016.]
Other Pathways Donate Electrons to the Respiratory Chain via Ubiquinone
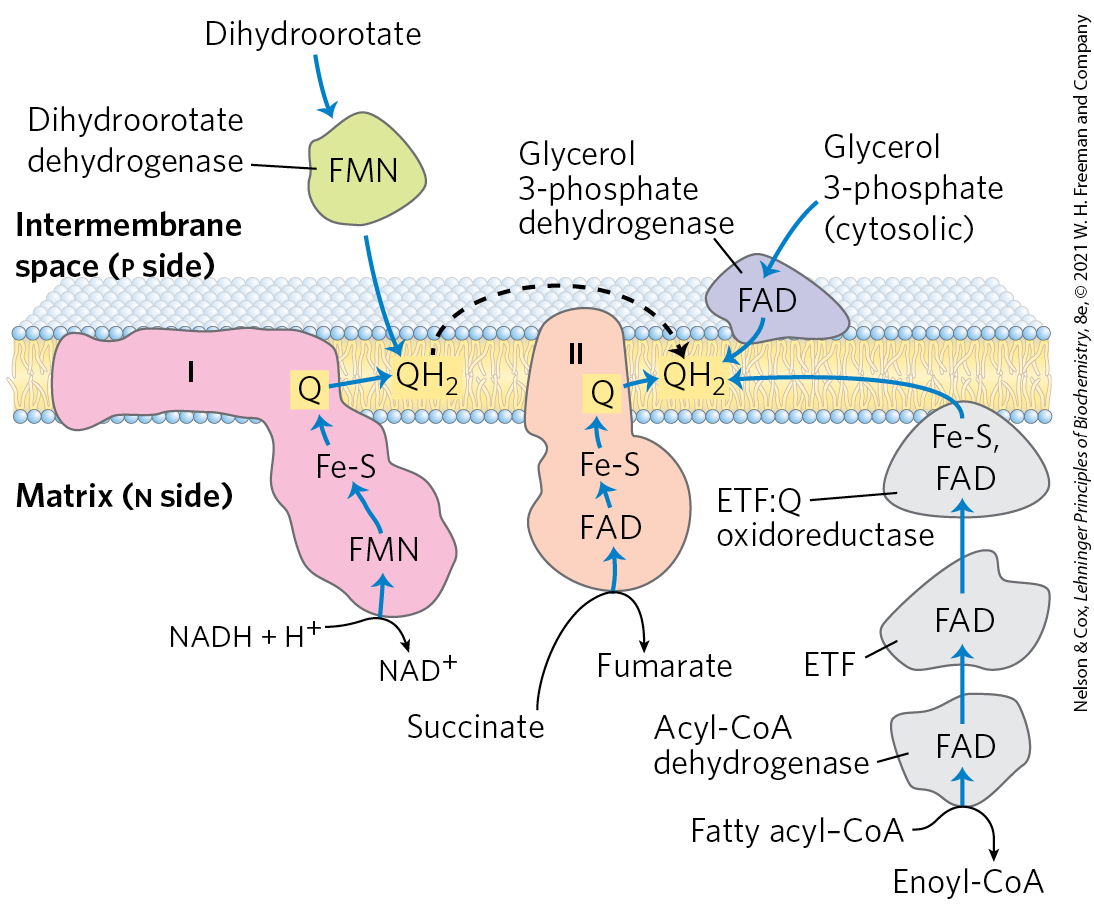
Several other electron-transfer reactions can reduce ubiquinone in the inner mitochondrial membrane (Fig. 19-15). In the first step of the β oxidation of fatty acyl–CoA, catalyzed by the flavoprotein acyl-CoA dehydrogenase (see Fig. 17-8), electrons pass from the substrate to the FAD of the dehydrogenase, then to electron-transferring flavoprotein (ETF). ETF passes its electrons to ETF: ubiquinone oxidoreductase, which reduces Q in the inner mitochondrial membrane to . Glycerol 3-phosphate, formed either from glycerol released by triacylglycerol breakdown or from the reduction of dihydroxyacetone phosphate from glycolysis, is oxidized by glycerol 3-phosphate dehydrogenase (see Fig. 17-4), a flavoprotein located on the outer face of the inner mitochondrial membrane. The electron acceptor in this reaction is Q; the produced enters the pool of in the membrane. The important role of glycerol 3-phosphate dehydrogenase in shuttling reducing equivalents from cytosolic NADH into the mitochondrial matrix is described in Section 19.2 (see Fig. 19-32). Dihydroorotate dehydrogenase, which acts in the synthesis of pyrimidines (see Fig. 22-38), is also on the outside of the inner mitochondrial membrane and donates electrons to Q in the respiratory chain. The reduced passes its electrons through Complex III and ultimately to .
FIGURE 19-15 Paths of electron transfer to ubiquinone in the respiratory chain. Electrons from NADH in the matrix pass through the FMN of a flavoprotein (NADH dehydrogenase) to a series of Fe-S centers (in Complex I) and then to Q. Electrons from succinate oxidation in the citric acid cycle pass through a flavoprotein with several Fe-S centers (Complex II) on the way to Q. Acyl-CoA dehydrogenase, the first enzyme of fatty acid β oxidation, transfers electrons to electron-transferring flavoprotein (ETF), from which they pass to Q via ETF:ubiquinone oxidoreductase. Dihydroorotate, an intermediate in the biosynthetic pathway to pyrimidine nucleotides, donates two electrons to Q through a flavoprotein (dihydroorotate dehydrogenase). And glycerol 3-phosphate, an intermediate of glycolysis in the cytosol, donates electrons to a flavoprotein (glycerol 3-phosphate dehydrogenase) on the outer face of the inner mitochondrial membrane, from which they pass to freely diffuses through the membrane (black dashed arrow), and can interact with several additional complexes.
The Energy of Electron Transfer Is Efficiently Conserved in a Proton Gradient
The transfer of two electrons from NADH through the respiratory chain to molecular oxygen can be summarized as
(19-5)
This net reaction is highly exergonic. For the redox pair , is , and for the pair , is . The for this reaction is therefore 1.14 V, and the standard free-energy change (see Eqn 13-7, p. 492) is
(19-6)
This standard free-energy change is based on the assumption of equal concentrations (1 m) of NADH and . In actively respiring mitochondria, the actions of many dehydrogenases keep the actual ratio well above unity, and the real free-energy change for the reaction shown in Equation 19-5 is therefore substantially greater (more negative) than . A similar calculation for the oxidation of succinate shows that electron transfer from succinate to has a smaller, but still negative, standard free-energy change of about .
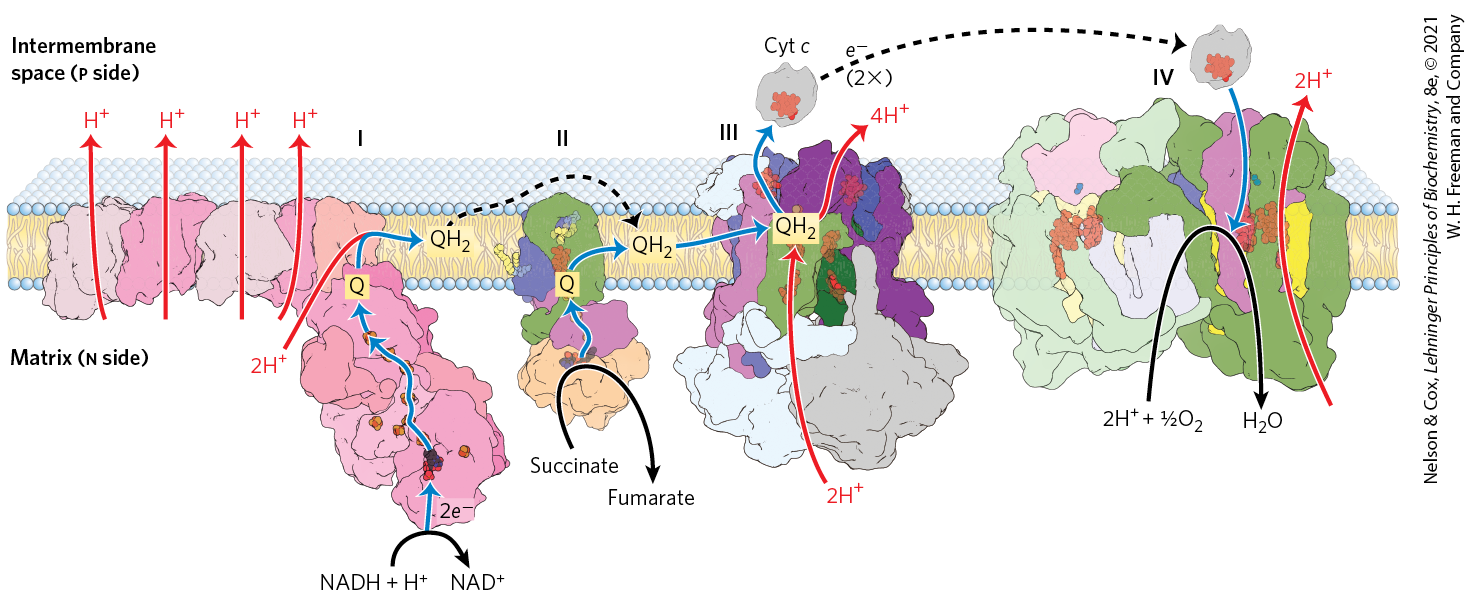
Much of this energy is used to pump protons out of the matrix. For each pair of electrons transferred to , four protons are pumped out by Complex I, four by Complex III, and two by Complex IV (Fig. 19-16). The vectorial equation for the process is therefore
(19-7)
FIGURE 19-16 Summary of the flow of electrons and protons through the four complexes of the respiratory chain. Electrons reach Q through Complexes I and II (as well as through several other paths shown in Fig. 19-15). Reduced Q serves as a mobile carrier of electrons and protons. It passes electrons to Complex III, which passes them to another mobile connecting link, cytochrome c. Complex IV then transfers electrons from reduced cytochrome c to . Electron flow through Complexes I, III, and IV is accompanied by proton efflux from the matrix into the intermembrane space. In bovine heart, the approximate ratios of Complexes I:II:III:IV are 1.1:1.3:3.0:6.7. Broken lines indicate the diffusion of Q in the plane of the inner membrane, and of cytochrome c through the intermembrane space. [Data from Complex I: PDB ID 4HEA, R. Baradaran et al., Nature 494:443, 2013; Complex II: PDB ID 1ZOY, F. Sun et al., Cell 121:1043, 2005; Complex III: PDB ID 1BGY, S. Iwata et al., Science 281:64, 1998; cytochrome c: PDB ID 1HRC, G. W. Bushnell et al., J. Mol. Biol. 214:585, 1990; Complex IV: PDB ID 1OCC, T. Tsukihara et al., Science 272:1136, 1996.]
The electrochemical energy inherent in this difference in proton concentration and separation of charge represents a temporary conservation of much of the energy of electron transfer. The energy stored in such a gradient, termed the proton-motive force, has two components: (1) the chemical potential energy due to the difference in concentration of a chemical species in the two regions separated by the membrane, and (2) the electrical potential energy that results from the separation of charge when a proton moves across the membrane without a counterion (Fig. 19-17).
FIGURE 19-17 Proton-motive force. The inner mitochondrial membrane separates two compartments of different , resulting in differences in chemical concentration and charge distribution across the membrane. The net effect is the proton-motive force , which can be calculated as shown here.
As we saw in Chapter 11, the free-energy change for the creation of an electrochemical gradient by an ion pump is
(19-8)
where and are the concentrations of an ion in two regions, and ; Z is the absolute value of its electrical charge (1 for a proton); and is the transmembrane difference in electrical potential, measured in volts.
In actively respiring mitochondria, the measured is 0.15 to 0.20 V, and the pH of the matrix is about 0.75 units more alkaline than that of the intermembrane space.
WORKED EXAMPLE 19-1 Energetics of Electron Transfer
Calculate the amount of energy conserved in the proton gradient across the inner mitochondrial membrane per pair of electrons transferred through the respiratory chain from NADH to oxygen. Assume is 0.15 V and the pH difference is 0.75 unit at body temperature of .
SOLUTION:
Equation 19-9 gives the free-energy change when one mole of protons moves across the inner membrane. Substituting the values of the constants R and F, 310 K for T, and the measured values for (0.75 unit) and (0.15 V) in this equation gives (of protons). Because the transfer of two electrons from NADH to is accompanied by the outward pumping of 10 protons (Eqn 19-7), roughly 190 kJ (of the 220 kJ released by oxidation of 1 mol of NADH) is conserved in the proton gradient.
When protons flow spontaneously down their electrochemical gradient, energy is made available to do work. In mitochondria, chloroplasts, and aerobic bacteria, the electrochemical energy in the proton gradient drives the synthesis of ATP from ADP and . We return to the energetics and stoichiometry of ATP synthesis driven by the electrochemical potential of the proton gradient in Section 19.2.
Reactive Oxygen Species Are Generated during Oxidative Phosphorylation
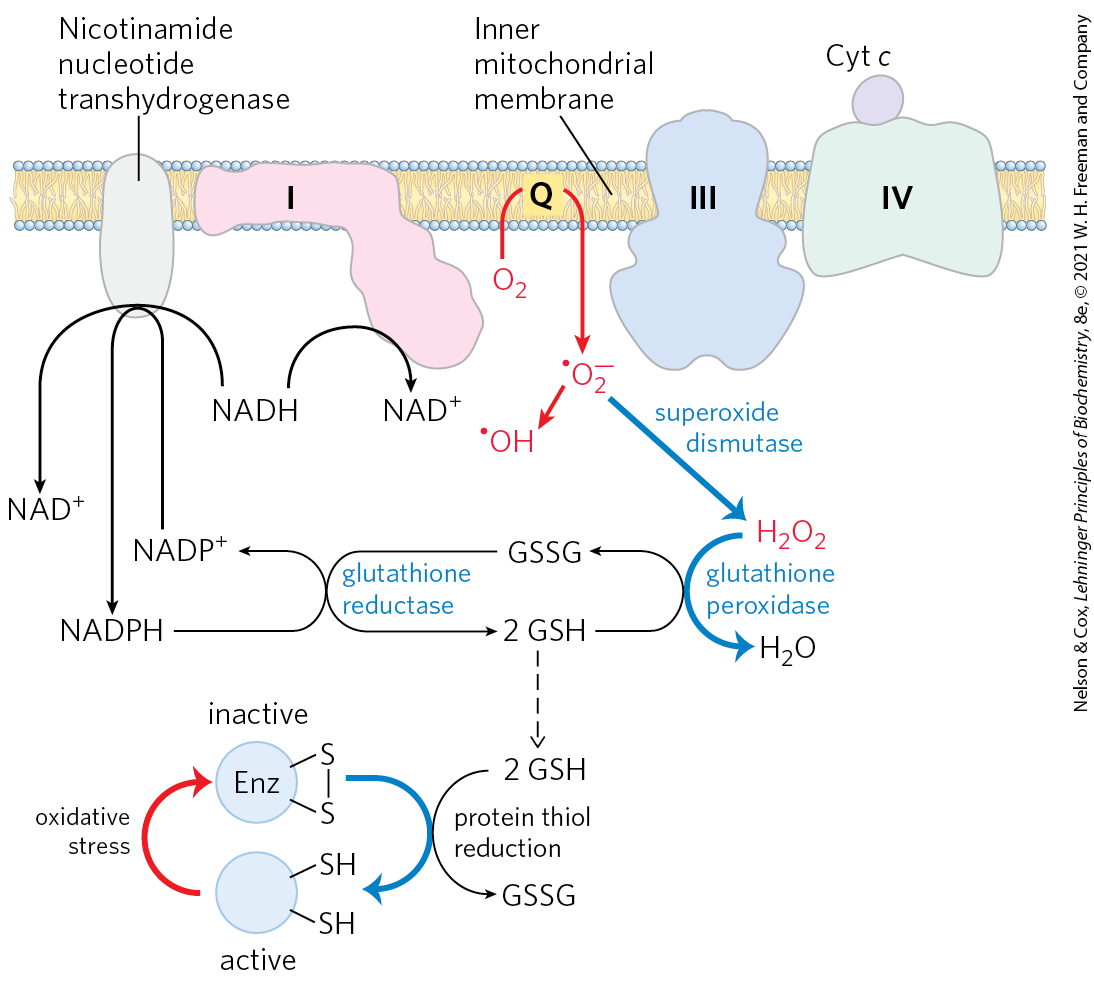
Several steps in the path of oxygen reduction in mitochondria have the potential to produce reactive oxygen species (superoxide, hydrogen peroxide, and hydroxyl radicals) that can damage cells. Some intermediates in the electron-transfer system, such as the partially reduced ubisemiquinone, can react directly with oxygen to form the superoxide radical as an intermediate. The radical is formed when a single electron is passed to in the reaction
Successive reduction of the superoxide radical with additional electrons produces , hydroxyl radicals , and finally . The very reactive hydroxyl radical can be especially damaging (Fig. 19-18).
FIGURE 19-18 ROS formation in mitochondria and mitochondrial defenses. When the rate of electron entry into the respiratory chain and the rate of electron transfer through the chain are mismatched, superoxide radical production increases at Complexes I and III as the partially reduced ubiquinone radical donates an electron to . Superoxide acts on aconitase, a 4Fe-4S protein (not shown), to release in turn will react with hydrogen peroxide (in a process called the Fenton reaction) to produce the highly reactive hydroxyl free radical . The reactions shown in blue defend the cell against the damaging effects of superoxide. Reduced glutathione (GSH; see Fig. 22-29) donates electrons for the reduction of and of the oxidized Cys residues of enzymes and other proteins; GSH is regenerated from the oxidized form (GSSG) by reduction with NADPH.
Reactive oxygen species (ROS) can wreak havoc, reacting with and damaging enzymes, membrane lipids, and nucleic acids. In actively respiring mitochondria, 0.2% to as much as 2% of the used in respiration forms — more than enough to have lethal effects unless the free radical is quickly disposed of. Factors that slow the flow of electrons through the respiratory chain increase the formation of superoxide, perhaps by prolonging the lifetime of generated in the Q cycle. The formation of ROS is favored when two conditions are met: (1) mitochondria are not making ATP (for lack of ADP or ) and therefore have a large proton-motive force and a high ratio, and (2) there is a high ratio in the matrix. In these situations, the mitochondrion is under oxidative stress — more electrons are available to enter the respiratory chain than can be immediately passed through to oxygen. When the supply of electron donors (NADH) is matched with that of electron acceptors, there is less oxidative stress, and ROS production is much reduced. Although overproduction of ROS is clearly detrimental, low levels of ROS are used by the cell as a signal reflecting the insufficient supply of oxygen (hypoxia), triggering metabolic adjustments (see Fig. 19-34).
To prevent oxidative damage by , cells have the enzyme superoxide dismutase, which catalyzes the reaction
The hydrogen peroxide thus generated is rendered harmless by glutathione peroxidase (Fig. 19-18). Glutathione reductase recycles the oxidized glutathione to its reduced form, using electrons from the NADPH generated by nicotinamide nucleotide transhydrogenase (in the mitochondrion) or by the pentose phosphate pathway (in the cytosol; see Fig. 14-30). Reduced glutathione also serves to keep protein sulfhydryl groups in their reduced state, preventing some of the deleterious effects of oxidative stress.
SUMMARY 19.1 The Mitochondrial Respiratory Chain
Chemiosmotic theory provides the intellectual framework for understanding many biological energy transductions, including oxidative phosphorylation and photophosphorylation. The energy of electron flow is conserved by the concomitant pumping of protons across the membrane, producing an electrochemical gradient, the proton-motive force.
In mitochondria, hydride ions removed from substrates (such as α-ketoglutarate and malate) by NAD-linked dehydrogenases donate electrons to the respiratory chain, which transfers the electrons to molecular , reducing it to .
Reducing equivalents from NADH are passed through a series of carrier types that include ubiquinone, Fe-S centers, and cytochromes.
Electron carriers are arranged in multiprotein complexes, which couple electron transfer to the generation of proton gradients. Complex I transfers electrons from NADH to ubiquinone. Complex II harvests electrons from the oxidation of succinate, also transferring them to ubiquinone. Ubiquinone diffuses through the inner mitochondrial membrane, and transfers the electrons to cytochrome b, the first carrier in Complex III. In this complex, electrons move first to an Fe-S center. The Fe-S center passes electrons, one at a time, through cytochrome c and into Complex IV, cytochrome oxidase. This copper-containing enzyme, which also contains cytochromes a and , accumulates electrons, then passes them to , reducing it to .
Complexes I, III, and IV form supercomplexes that facilitate electron flow between them.
Some electrons enter this chain of carriers through alternative paths. Electrons derived from the oxidation of fatty acids pass to ubiquinone via the electron-transferring flavoprotein. The oxidation of glycerol phosphate and of dihydroorotate also sends electrons into the respiratory chain at the level of .
Much of the free energy generated by electron transfer and the reduction of oxygen to form water is recovered and stored in the form of an electrochemical proton gradient across the mitochondrial inner membrane.
Potentially harmful reactive oxygen species produced in mitochondria are inactivated by a set of protective enzymes, including superoxide dismutase and glutathione peroxidase. Low levels of ROS serve as signals coordinating mitochondrial oxidative phosphorylation with other metabolic pathways.

 The mitochondrial matrix, enclosed by the inner membrane, contains the pyruvate dehydrogenase complex and the enzymes of the citric acid cycle, the fatty acid β-oxidation pathway, and the pathways of amino acid oxidation — all the pathways of fuel oxidation except glycolysis, which takes place in the cytosol. The selectively permeable inner mitochondrial membrane segregates the intermediates and enzymes of cytosolic metabolic pathways from those of metabolic processes occurring in the matrix. However, specific transporters carry pyruvate, fatty acids, and amino acids or their α-keto derivatives into the matrix for access to the machinery of the citric acid cycle. ADP and are specifically transported into the matrix as newly synthesized ATP is transported out. Mammalian mitochondria have about 1,200 proteins, according to current best estimates. The functions of up to 25% of these remain partly or entirely enigmatic.
The mitochondrial matrix, enclosed by the inner membrane, contains the pyruvate dehydrogenase complex and the enzymes of the citric acid cycle, the fatty acid β-oxidation pathway, and the pathways of amino acid oxidation — all the pathways of fuel oxidation except glycolysis, which takes place in the cytosol. The selectively permeable inner mitochondrial membrane segregates the intermediates and enzymes of cytosolic metabolic pathways from those of metabolic processes occurring in the matrix. However, specific transporters carry pyruvate, fatty acids, and amino acids or their α-keto derivatives into the matrix for access to the machinery of the citric acid cycle. ADP and are specifically transported into the matrix as newly synthesized ATP is transported out. Mammalian mitochondria have about 1,200 proteins, according to current best estimates. The functions of up to 25% of these remain partly or entirely enigmatic. During cell growth and division, mitochondria, like bacteria, divide by fission, and under some circumstances individual mitochondria fuse to form larger, more-extended structures. Stressful conditions, such as the presence of electron-transfer inhibitors or mutations in an electron carrier, trigger mitochondrial fission and sometimes mitophagy — the breakdown of mitochondria and recycling of the amino acids, nucleotides, and lipids released. As stress is relieved, small mitochondria fuse to form long, thin, tubular organelles.
During cell growth and division, mitochondria, like bacteria, divide by fission, and under some circumstances individual mitochondria fuse to form larger, more-extended structures. Stressful conditions, such as the presence of electron-transfer inhibitors or mutations in an electron carrier, trigger mitochondrial fission and sometimes mitophagy — the breakdown of mitochondria and recycling of the amino acids, nucleotides, and lipids released. As stress is relieved, small mitochondria fuse to form long, thin, tubular organelles. The oxidized flavin nucleotide can accept either one electron (yielding the semiquinone form) or two (yielding or ). Electron transfer occurs because the flavoprotein has a higher reduction potential than the compound oxidized. Recall that reduction potential is a quantitative measure of the relative tendency of a given chemical species to accept electrons in an oxidation-reduction reaction (
The oxidized flavin nucleotide can accept either one electron (yielding the semiquinone form) or two (yielding or ). Electron transfer occurs because the flavoprotein has a higher reduction potential than the compound oxidized. Recall that reduction potential is a quantitative measure of the relative tendency of a given chemical species to accept electrons in an oxidation-reduction reaction (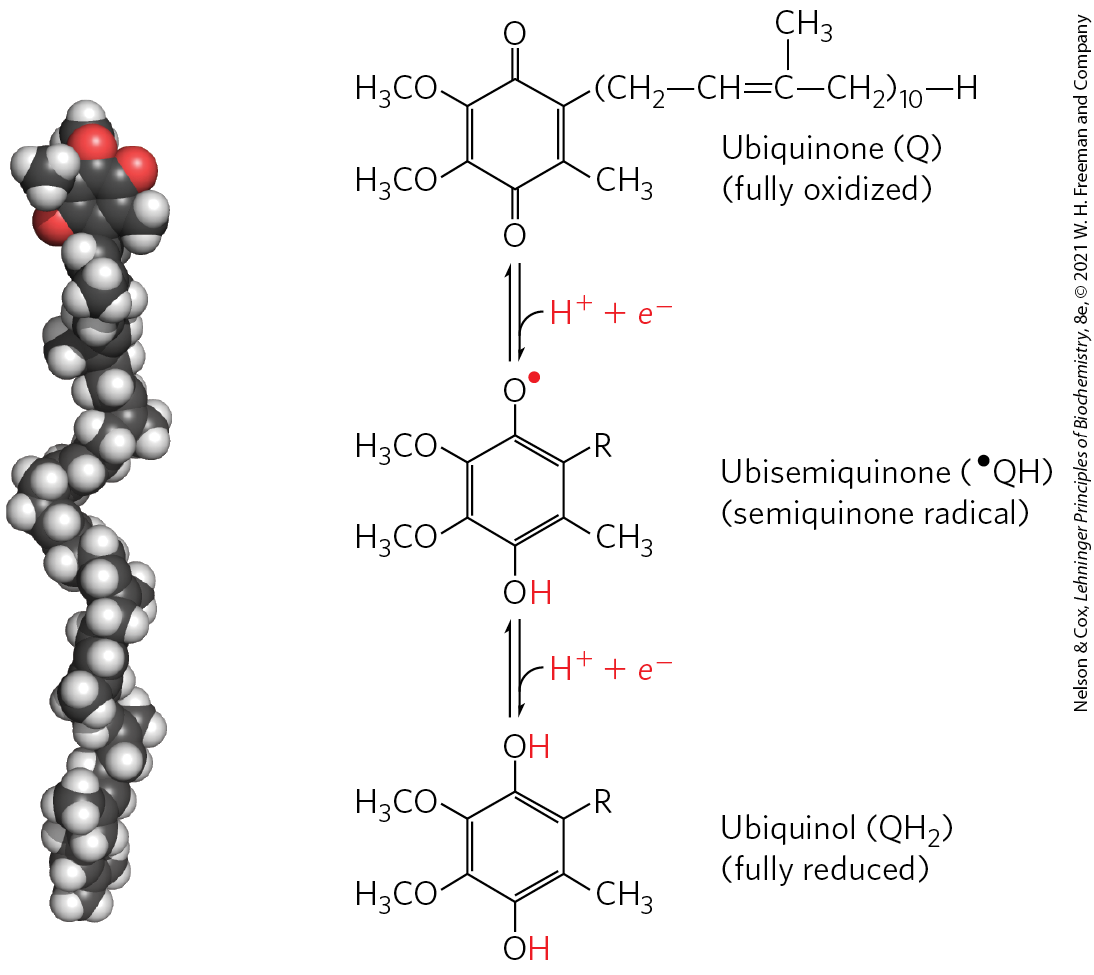

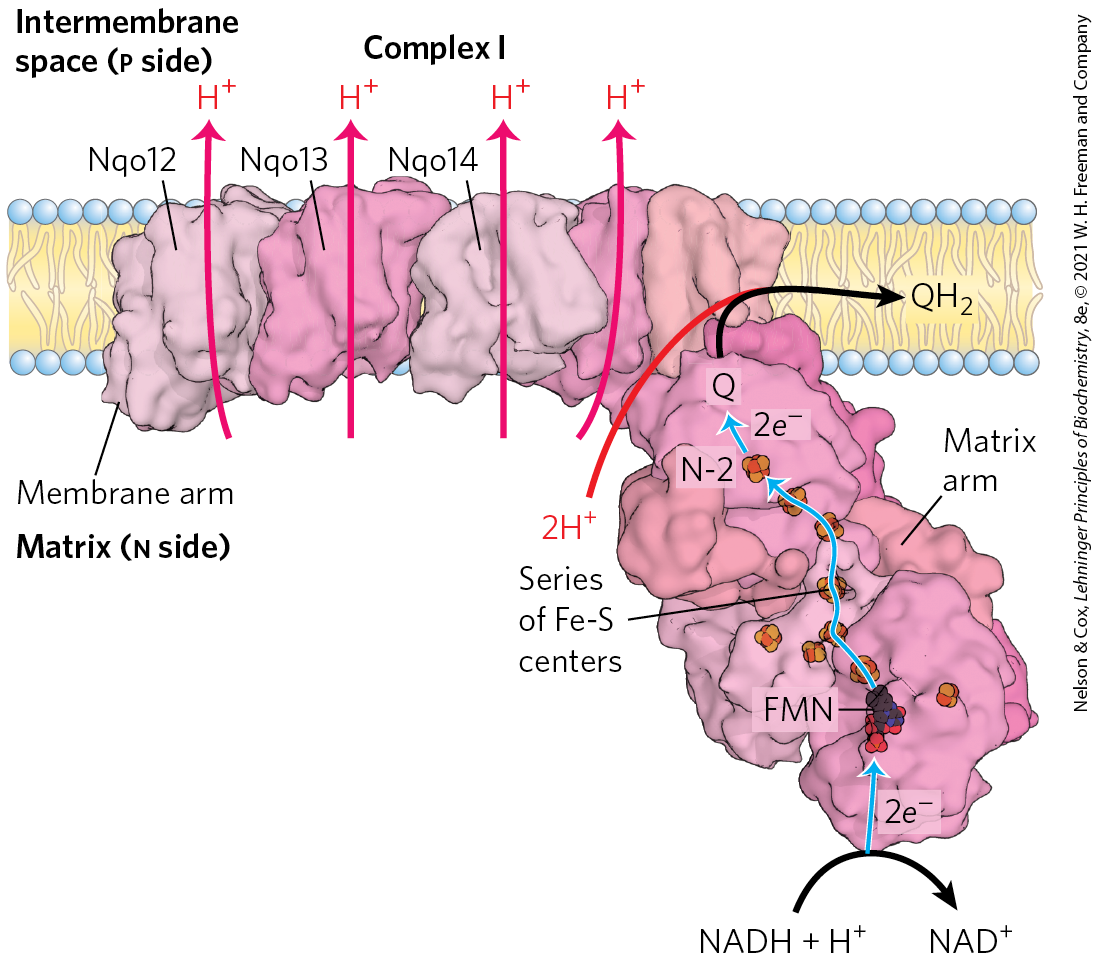
 Complex I is therefore a proton pump driven by the energy of electron transfer, and the reaction it catalyzes is
Complex I is therefore a proton pump driven by the energy of electron transfer, and the reaction it catalyzes is 
 The heme b of Complex II is apparently not in the direct path of electron transfer; it serves instead to reduce the frequency with which electrons “leak” out of the system, moving from succinate to molecular oxygen to produce the
The heme b of Complex II is apparently not in the direct path of electron transfer; it serves instead to reduce the frequency with which electrons “leak” out of the system, moving from succinate to molecular oxygen to produce the 
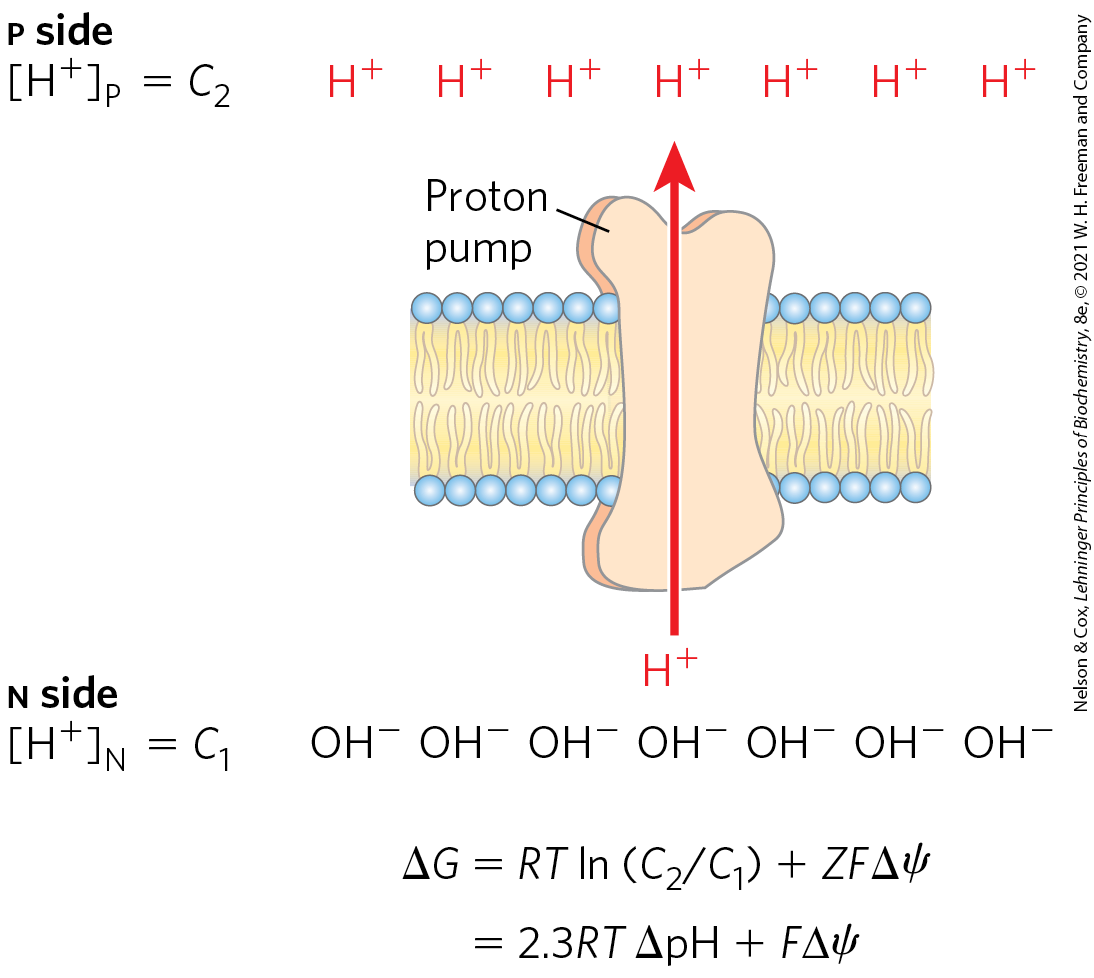
 Chemiosmotic theory provides the intellectual framework for understanding many biological energy transductions, including oxidative phosphorylation and photophosphorylation. The energy of electron flow is conserved by the concomitant pumping of protons across the membrane, producing an electrochemical gradient, the proton-motive force.
Chemiosmotic theory provides the intellectual framework for understanding many biological energy transductions, including oxidative phosphorylation and photophosphorylation. The energy of electron flow is conserved by the concomitant pumping of protons across the membrane, producing an electrochemical gradient, the proton-motive force.