16.2 Reactions of the Citric Acid Cycle
We are now ready to trace the process by which acetyl-CoA undergoes oxidation. This chemical transformation is carried out by the citric acid cycle, the first cyclic pathway we have encountered (Fig. 16-7). To begin a turn of the cycle, acetyl-CoA donates its acetyl group to the four-carbon compound oxaloacetate to form the six-carbon citrate. Citrate is then transformed into isocitrate, also a six-carbon molecule, which is dehydrogenated with loss of to yield the five-carbon compound α-ketoglutarate (also called 2-oxoglutarate). α-Ketoglutarate undergoes loss of a second molecule of and ultimately yields the four-carbon compound succinate. Succinate is enzymatically converted in three steps into the four-carbon oxaloacetate — which is then ready to react with another molecule of acetyl-CoA. In each turn of the cycle, one acetyl group (two carbons) enters as acetyl-CoA and two molecules of leave; one molecule of oxaloacetate is used to form citrate and one molecule of oxaloacetate is regenerated. No net removal of oxaloacetate occurs; one molecule of oxaloacetate can theoretically bring about oxidation of an infinite number of acetyl groups, effectively acting catalytically; the steady-state concentration of oxaloacetate is very low (micromolar). Four of the eight steps in this process are oxidations, in which the energy of oxidation is very efficiently conserved in the form of the reduced coenzymes NADH and . These two carriers donate their electrons to the respiratory chain where electron flow drives ATP synthesis.
 one molecule of oxaloacetate can theoretically bring about oxidation of an infinite number of acetyl groups, effectively acting catalytically; the steady-state concentration of oxaloacetate is very low (micromolar). Four of the eight steps in this process are oxidations, in which the energy of oxidation is very efficiently conserved in the form of the reduced coenzymes NADH and . These two carriers donate their electrons to the respiratory chain where electron flow drives ATP synthesis.
one molecule of oxaloacetate can theoretically bring about oxidation of an infinite number of acetyl groups, effectively acting catalytically; the steady-state concentration of oxaloacetate is very low (micromolar). Four of the eight steps in this process are oxidations, in which the energy of oxidation is very efficiently conserved in the form of the reduced coenzymes NADH and . These two carriers donate their electrons to the respiratory chain where electron flow drives ATP synthesis.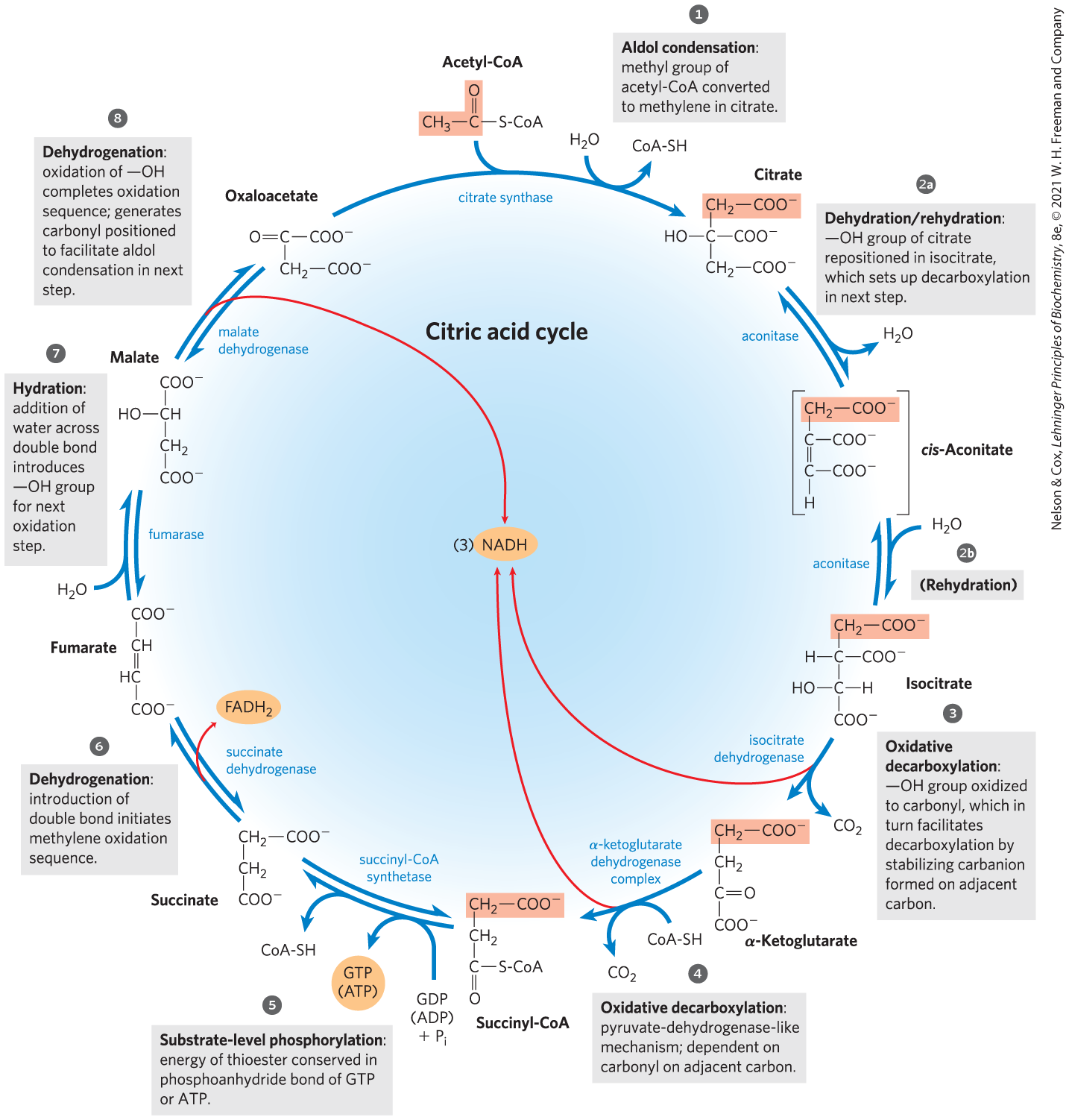
FIGURE 16-7 Reactions of the citric acid cycle. The carbon atoms shaded in pink are those derived from the acetate of acetyl-CoA in the first turn of the cycle; these are not the carbons released as in the first turn. Note that in succinate and fumarate, the two-carbon group derived from acetate can no longer be specifically denoted; because succinate and fumarate are symmetric molecules, C-1 and C-2 are indistinguishable from C-4 and C-3. The red arrows show where energy is conserved by electron transfer to FAD or , forming or . Steps , , and are essentially irreversible in the cell; all other steps are reversible. The nucleoside triphosphate product of step may be either ATP or GTP, depending on which succinyl-CoA synthetase isozyme is the catalyst.
 ,
,  , and
, and  are essentially irreversible in the cell; all other steps are reversible. The nucleoside triphosphate product of step
are essentially irreversible in the cell; all other steps are reversible. The nucleoside triphosphate product of step  may be either ATP or GTP, depending on which succinyl-CoA synthetase isozyme is the catalyst.
may be either ATP or GTP, depending on which succinyl-CoA synthetase isozyme is the catalyst.Although the citric acid cycle is central to energy-yielding metabolism, its role is not limited to energy conservation. Four- and five-carbon intermediates of the cycle serve as precursors for a wide variety of products. To replace intermediates removed for this purpose, cells employ anaplerotic (replenishing) reactions, which are described below.
Eugene Kennedy and Albert Lehninger showed in 1948 that, in eukaryotes, the entire set of reactions of the citric acid cycle takes place in mitochondria. Isolated mitochondria were found to contain not only all the enzymes and coenzymes required for the citric acid cycle, but also all the enzymes and proteins necessary for the last stage of respiration — electron transfer and ATP synthesis by oxidative phosphorylation. As we shall see in later chapters, mitochondria also contain the enzymes for the oxidation of fatty acids and some amino acids to acetyl-CoA, and the oxidative degradation of other amino acids to α-ketoglutarate, succinyl-CoA, or oxaloacetate. Thus, in nonphotosynthetic eukaryotes, the mitochondrion is the site of most energy-yielding oxidative reactions and of the coupled synthesis of ATP. In photosynthetic eukaryotes, mitochondria are the major site of ATP production in the dark, but in daylight, chloroplasts produce most of the organism’s ATP. In most bacteria, the enzymes of the citric acid cycle are in the cytosol, and the plasma membrane plays a role analogous to that of the inner mitochondrial membrane in ATP synthesis (Chapter 19).
The Sequence of Reactions in the Citric Acid Cycle Makes Chemical Sense
Acetyl-CoA produced in the breakdown of carbohydrates, fats, and proteins must be completely oxidized to if the maximum potential energy is to be extracted from these fuels. However, it is not biochemically feasible to directly oxidize acetate (or acetyl-CoA) to . Decarboxylation of this two-carbon acid would yield and methane . Methane is chemically rather stable, and except for certain methanotrophic bacteria that grow in methane-rich niches, organisms do not have the cofactors and enzymes needed to oxidize methane. Methylene groups , however, are readily metabolized by enzyme systems present in most organisms. In typical oxidation sequences, two adjacent methylene groups are involved, at least one of which is adjacent to a carbonyl group. As we noted in Chapter 13 (p. 473), carbonyl groups are particularly important in the chemical transformations of metabolic pathways. The carbon of the carbonyl group has a partial positive charge due to the electron-withdrawing property of the carbonyl oxygen and is therefore an electrophilic center. A carbonyl group can facilitate the formation of a carbanion on an adjoining carbon by delocalizing the carbanion’s negative charge. We see an example of the oxidation of a methylene group in the citric acid cycle, as succinate is oxidized (steps to in Fig. 16-7) to form a carbonyl (in oxaloacetate) that is more chemically reactive than either a methylene group or methane.
 to
to  in
in In short, if acetyl-CoA is to be oxidized efficiently, the methyl group of the acetyl-CoA must be attached to something. The first step of the citric acid cycle — the condensation of acetyl-CoA with oxaloacetate — neatly solves the problem of the unreactive methyl group. The carbonyl of oxaloacetate acts as an electrophilic center, which is attacked by the methyl carbon of acetyl-CoA in an aldol condensation (p. 473) to form citrate (step in Fig. 16-7). The methyl group of acetate has been converted into a methylene in citric acid. This tricarboxylic acid then readily undergoes a series of oxidations that eliminate two carbons as . Note that all steps featuring the breakage or formation of carbon–carbon bonds (steps , , and ) rely on properly positioned carbonyl groups. As in all metabolic pathways, there is a chemical logic to the sequence of steps in the citric acid cycle: each step either involves an energy-conserving oxidation or is a necessary prelude to the oxidation, placing functional groups in position to facilitate oxidation or oxidative decarboxylation. As you learn the steps of the cycle, keep in mind the chemical rationale for each; it will make the process easier to understand and remember.
The Citric Acid Cycle Has Eight Steps
In examining the eight successive reaction steps of the citric acid cycle, we place special emphasis on the chemical transformations taking place as citrate formed from acetyl-CoA and oxaloacetate is oxidized to yield and the energy of this oxidation is conserved in the reduced coenzymes NADH and .
Formation of Citrate
The first reaction of the cycle is the condensation of acetyl-CoA with oxaloacetate to form citrate, catalyzed by citrate synthase.
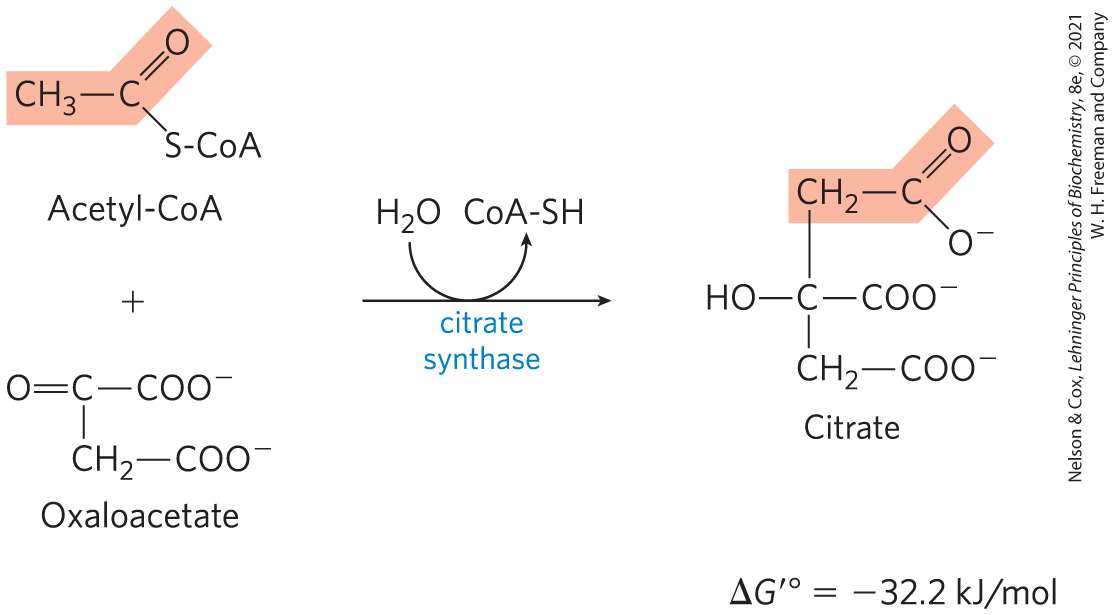
In this reaction, the methyl carbon of the acetyl group is joined to the carbonyl group (C-2) of oxaloacetate. Citroyl-CoA is a transient intermediate formed on the active site of the enzyme (see Fig. 16-9). It undergoes hydrolysis to free CoA and citrate, which are released from the active site. The hydrolysis of this high-energy thioester intermediate makes the forward reaction highly exergonic. The large, negative standard free-energy change of the forward citrate synthase reaction is essential to the operation of the cycle because the concentration of oxaloacetate is normally very low (micromolar). The CoA liberated in this reaction is recycled to participate in the oxidative decarboxylation of another molecule of pyruvate by the PDH complex.
Citrate synthase from mitochondria is a homodimer (Fig. 16-8). Each subunit is a single polypeptide with two domains, one large and rigid, the other smaller and more flexible, with the active site between them. Oxaloacetate, the first substrate to bind to the enzyme, induces a large conformational change in the flexible domain, creating a binding site for the second substrate, acetyl-CoA. When citroyl-CoA has formed in the enzyme active site, another conformational change brings about thioester hydrolysis, releasing CoA-SH. This induced fit of the enzyme first to its substrate and then to its reaction intermediate decreases the likelihood of premature and unproductive cleavage of the thioester bond of acetyl-CoA. Kinetic studies of the enzyme are consistent with this ordered bisubstrate mechanism (see Fig. 6-15). The reaction catalyzed by citrate synthase is essentially an aldol condensation (p. 473), involving a thioester (acetyl-CoA) and a ketone (oxaloacetate) (Fig. 16-9).
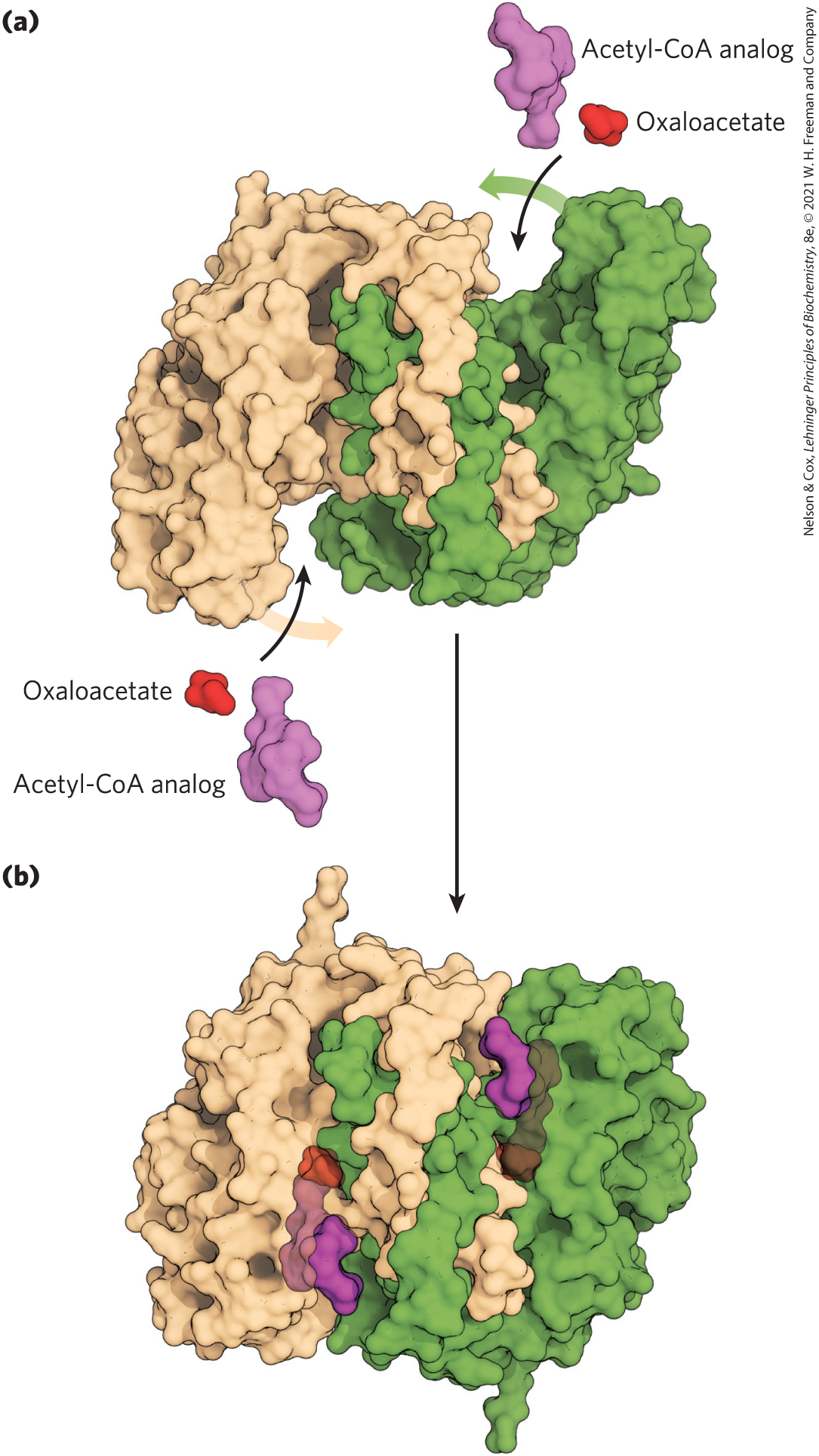
FIGURE 16-8 Structure of citrate synthase. The flexible domain of each subunit undergoes a large conformational change on binding oxaloacetate, creating a binding site for acetyl-CoA. (a) Open form of the enzyme alone; (b) closed form with bound oxaloacetate and a stable analog of acetyl-CoA (carboxymethyl-CoA). In these representations one subunit is colored tan and one green. [Data from (a) PDB ID 5CSC, D.-I. Liao et al., Biochemistry 30:6031, 1991;]
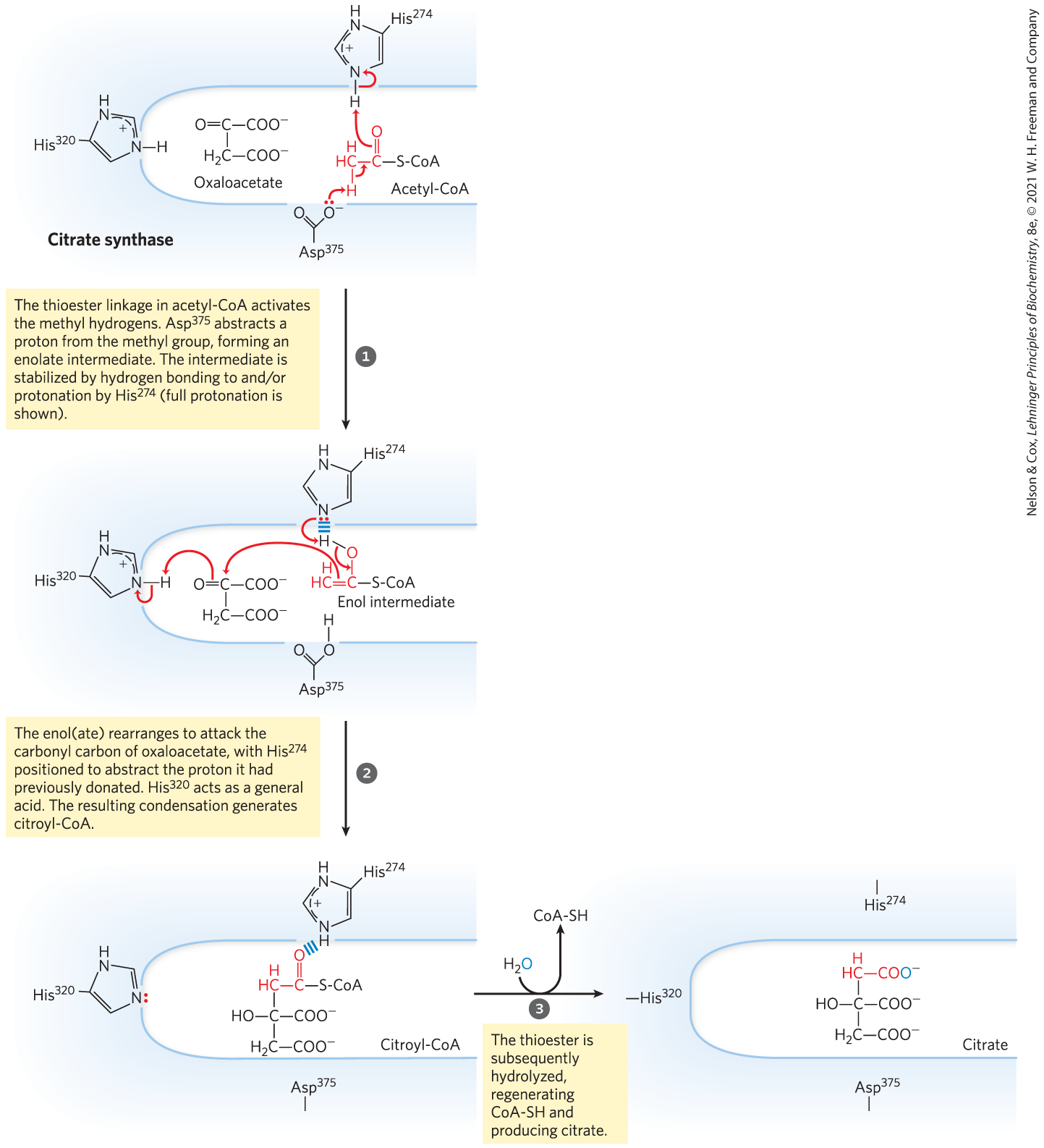
MECHANISM FIGURE 16-9 Citrate synthase. In the citrate synthase reaction in mammals, oxaloacetate binds first, in a strictly ordered reaction sequence. This binding triggers a conformation change that opens up the binding site for acetyl-CoA. Oxaloacetate is specifically oriented in the active site of citrate synthase by interaction of its two carboxylates with two positively charged Arg residues (not shown here). [Information from S. J. Remington, Curr. Opin. Struct. Biol. 2:730, 1992.]
Formation of Isocitrate via cis-Aconitate
 Formation of Isocitrate via cis-Aconitate
Formation of Isocitrate via cis-AconitateThe enzyme aconitase (more formally, aconitate hydratase) catalyzes the reversible transformation of citrate to isocitrate, through the intermediary formation of the tricarboxylic acid cis-aconitate, which normally does not dissociate from the active site. Aconitase can promote the reversible addition of to the double bond of enzyme-bound cis-aconitate in two different ways, one leading to citrate and the other to isocitrate:
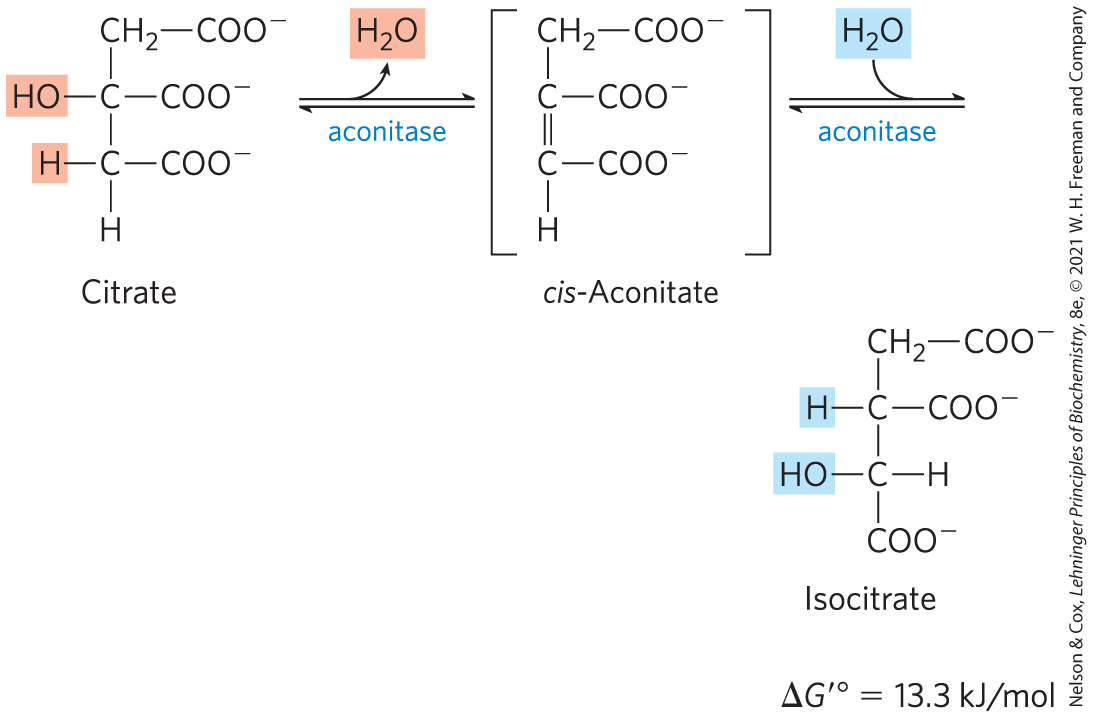
Although the equilibrium mixture at pH 7.4 and 25 °C contains less than 10% isocitrate, in the cell the reaction is pulled to the right because isocitrate is immediately consumed in the next step of the cycle, lowering its steady-state concentration. Aconitase contains an iron-sulfur center (Fig. 16-10), which acts both in the binding of the substrate at the active site and in the catalytic addition or removal of . In iron-depleted cells, acon-itase loses its iron-sulfur center and acquires a new role in the regulation of iron homeostasis. Aconitase is one of many enzymes known to “moonlight” in a second role (Box 16-1).
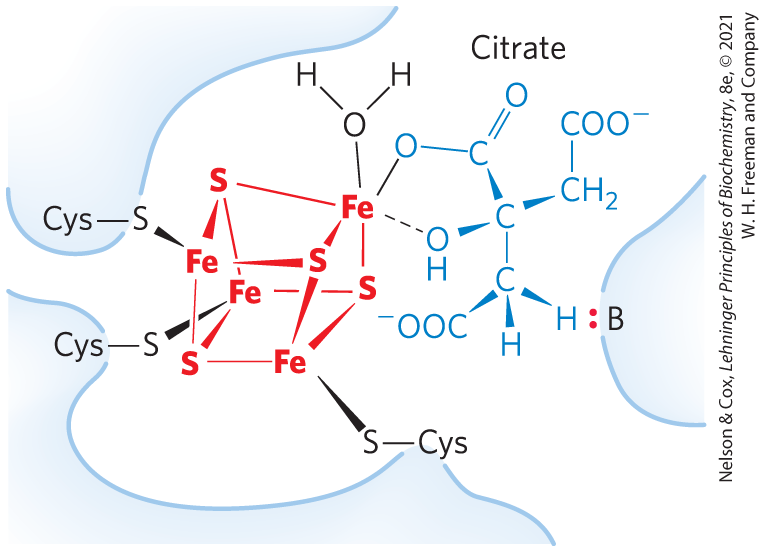
FIGURE 16-10 Iron-sulfur center in aconitase. The iron-sulfur center is in red, the citrate molecule in blue. Three Cys residues of the enzyme bind three iron atoms; the fourth iron is bound to one of the carboxyl groups of citrate and also interacts noncovalently with a hydroxyl group of citrate (dashed bond). A basic residue (:B) in the enzyme helps to position the citrate in the active site. The iron-sulfur center acts in both substrate binding and catalysis. The general properties of iron-sulfur proteins are discussed in Chapter 19.
Oxidation of Isocitrate to α-Ketoglutarate and
In the next step, isocitrate dehydrogenase catalyzes oxidative decarboxylation of isocitrate to form α-ketoglutarate (Fig. 16-11). in the active site interacts with the carbonyl group of the intermediate oxalosuccinate, which is formed transiently but does not leave the binding site until decarboxylation converts it to α-ketoglutarate. also stabilizes the enol formed transiently by decarboxylation.
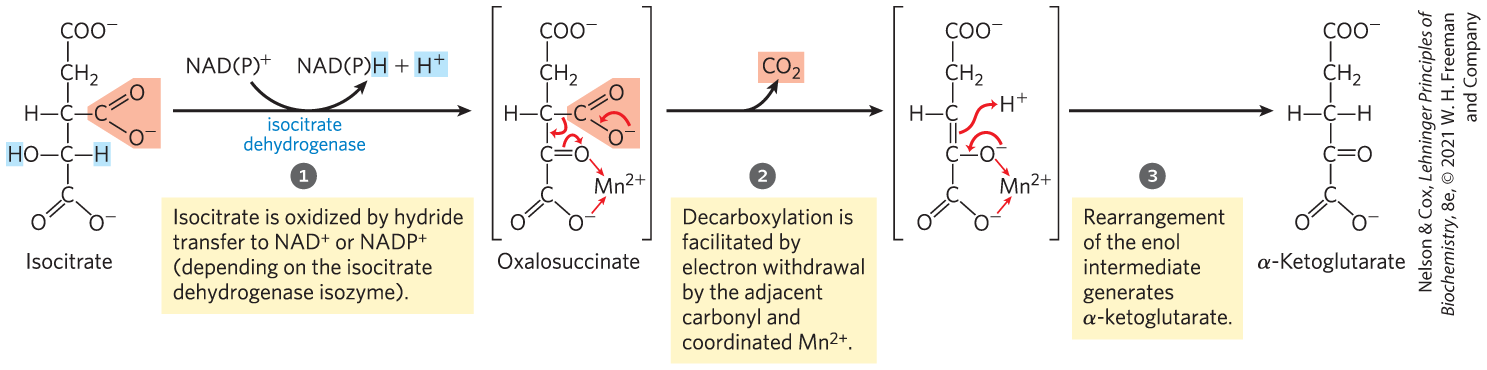
MECHANISM FIGURE 16-11 Isocitrate dehydrogenase. In this reaction, the substrate, isocitrate, loses one carbon by oxidative decarboxylation.
There are two different forms of isocitrate dehydrogenase in all cells, one requiring as electron acceptor and the other requiring . The overall reactions are otherwise identical. In eukaryotic cells, the NAD-dependent enzyme occurs in the mitochondrial matrix and serves in the citric acid cycle. The main function of the NADP-dependent enzyme, found in both the mitochondrial matrix and the cytosol, is the generation of NADPH, which is essential for reductive anabolic pathways such as fatty acid and sterol synthesis.
Oxidation of α-Ketoglutarate to Succinyl-CoA and
The next step is another oxidative decarboxylation, in which α-ketoglutarate is converted to succinyl-CoA and by the action of the α-ketoglutarate dehydrogenase complex; serves as electron acceptor and CoA as the carrier of the succinyl group. The energy of oxidation of α-ketoglutarate is conserved in the formation of the thioester bond of succinyl-CoA:
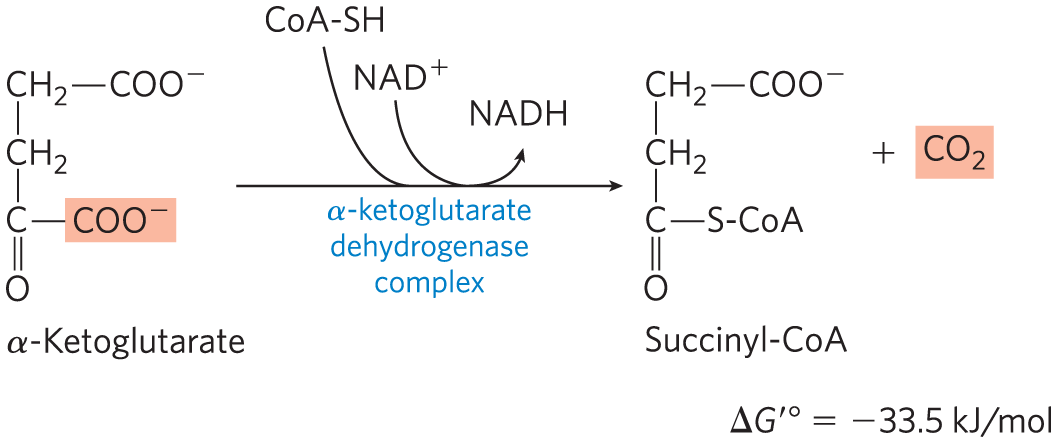
This reaction is virtually identical to the pyruvate dehydrogenase reaction discussed above and to the reaction sequence responsible for the breakdown of branched-chain amino acids (Fig. 16-12). The α-ketoglutarate dehydrogenase complex closely resembles the PDH complex and the complex that degrades branched-chain α-keto acids in both structure and function. All three have homologous and components, identical components, enzyme-bound TPP and lipoate, coenzyme A, FAD, and NAD. These related enzymes can employ the same subunit because the substrate for — a reduced lipoate — is the same for both complexes. They are certainly derived from a common evolutionary ancestor by gene duplication and subsequent divergent evolution, as described in Figure 1-32.

FIGURE 16-12 A conserved mechanism for oxidative decarboxylation. The pathways shown employ the same five cofactors (thiamine pyrophosphate, coenzyme A, lipoate, FAD, and ), closely similar multienzyme complexes, and the same enzymatic mechanism to carry out oxidative decarboxylations of pyruvate (by the pyruvate dehydrogenase complex), α-ketoglutarate (in the citric acid cycle), and the carbon skeletons of the three branched-chain amino acids, isoleucine (shown here), leucine, and valine.
Conversion of Succinyl-CoA to Succinate
Succinyl-CoA, like acetyl-CoA, has a thioester bond with a strongly negative standard free energy of hydrolysis . In the next step of the citric acid cycle, energy released in the breakage of this bond is used to drive the synthesis of a phosphoanhydride bond in either GTP or ATP, with a net of only –2.9 kJ/mol. Succinate is formed in the process:
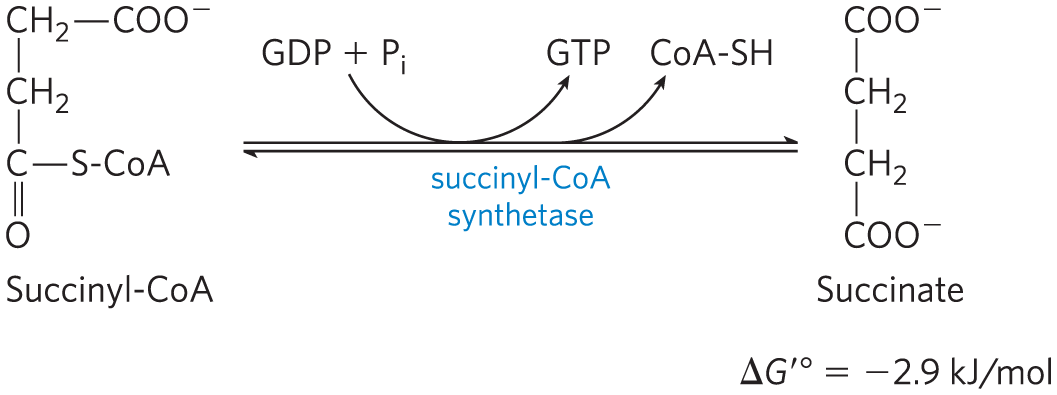
The enzyme that catalyzes this reversible reaction is called succinyl-CoA synthetase or succinic thiokinase; both names indicate the participation of a nucleoside triphosphate in the reaction.
This energy-conserving reaction involves an intermediate step in which the enzyme molecule itself becomes phosphorylated at a His residue in the active site (Fig. 16-13a). This phosphoryl group, which has a high group transfer potential, is transferred to ADP (or GDP) to form ATP (or GTP). Animal cells have two isozymes of succinyl-CoA synthetase, one specific for ADP and the other for GDP. The enzyme has two subunits, , which has the –His residue and the binding site for CoA, and , which confers specificity for either ADP or GDP. The active site is at the interface between subunits. The crystal structure of succinyl-CoA synthetase reveals two “power helices” (one from each subunit), oriented so that their electric dipoles situate partial positive charges close to the negatively charged –His (Fig. 16-13b), stabilizing the phosphoenzyme intermediate. (Recall the similar role of helix dipoles in stabilizing ions in the channel; see Fig. 11-45.)
 –His residue and the binding site for CoA, and , which confers specificity for either ADP or GDP. The active site is at the interface between subunits. The crystal structure of succinyl-CoA synthetase reveals two “power helices” (one from each subunit), oriented so that their electric dipoles situate partial positive charges close to the negatively charged
–His residue and the binding site for CoA, and , which confers specificity for either ADP or GDP. The active site is at the interface between subunits. The crystal structure of succinyl-CoA synthetase reveals two “power helices” (one from each subunit), oriented so that their electric dipoles situate partial positive charges close to the negatively charged 
FIGURE 16-13 The succinyl-CoA synthetase reaction. (a) In step a phosphoryl group replaces the CoA of succinyl-CoA bound to the enzyme, forming a high-energy acyl phosphate. In step the succinyl phosphate donates its phosphoryl group to a His residue of the enzyme, forming a high-energy phosphohistidyl enzyme. In step the phosphoryl group is transferred from the His residue to the terminal phosphate of GDP (or ADP), forming GTP (or ATP). (b) Active site of succinyl-CoA synthetase of Escherichia coli. The active site includes part of both the α (blue) and the β (brown) subunits. The power helices (blue, brown) place the partial positive charges of the helix dipole near the phosphate group of in the α chain, stabilizing the phosphohistidyl enzyme. The bacterial and mammalian enzymes have similar amino acid sequences and three-dimensional structures. [Data from PDB ID 1SCU, W. T. Wolodko et al., J. Biol. Chem. 269:10,883, 1994.]
 in the α chain, stabilizing the phosphohistidyl enzyme. The bacterial and mammalian enzymes have similar amino acid sequences and three-dimensional structures. [Data from PDB ID 1SCU, W. T. Wolodko et al., J. Biol. Chem. 269:10,883, 1994.]
in the α chain, stabilizing the phosphohistidyl enzyme. The bacterial and mammalian enzymes have similar amino acid sequences and three-dimensional structures. [Data from PDB ID 1SCU, W. T. Wolodko et al., J. Biol. Chem. 269:10,883, 1994.]The formation of ATP (or GTP) at the expense of the energy released by the oxidative decarboxylation of α-ketoglutarate is a substrate-level phosphorylation, like the synthesis of ATP in the glycolytic reactions catalyzed by phosphoglycerate kinase and pyruvate kinase (see Fig. 14-2). The GTP formed by succinyl-CoA synthetase can donate its terminal phosphoryl group to ADP to form ATP, in a reversible reaction catalyzed by nucleoside diphosphate kinase (p. 487):
Thus the net result of the activity of either isozyme of succinyl-CoA synthetase is the conservation of energy as ATP. There is no change in free energy for the nucleoside diphosphate kinase reaction; ATP and GTP are energetically equivalent.
Oxidation of Succinate to Fumarate
The succinate formed from succinyl-CoA is oxidized to fumarate by the flavoprotein succinate dehydrogenase:
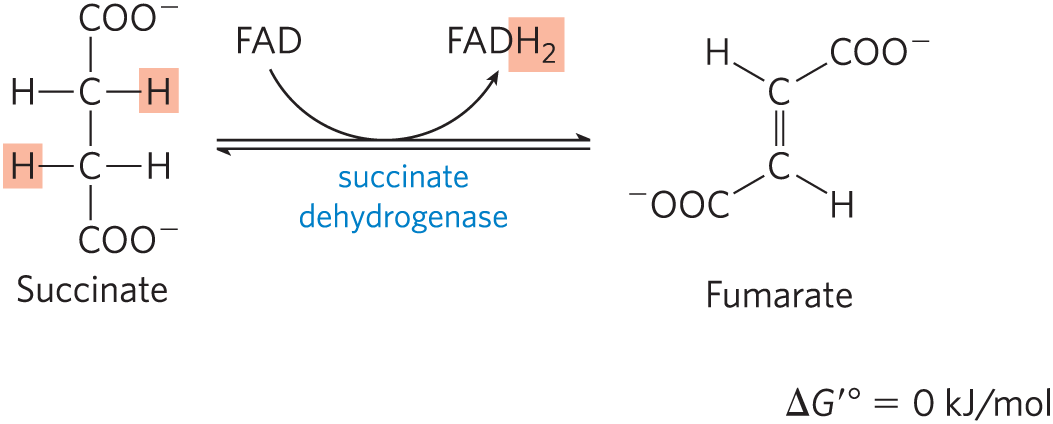
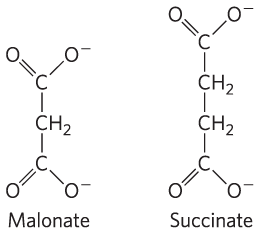
In eukaryotes, succinate dehydrogenase is an integral protein of the mitochondrial inner membrane; in bacteria, of the plasma membrane. The enzyme contains three different iron-sulfur clusters and one molecule of covalently bound FAD (see Fig. 19-9). Electrons pass from succinate through the FAD and iron-sulfur centers before entering the chain of electron carriers in the mitochondrial inner membrane (the plasma membrane in bacteria). Electron flow from succinate through these carriers to the final electron acceptor, , is coupled to the synthesis of about 1.5 ATP molecules per pair of electrons (respiration-linked phosphorylation). Malonate, an analog of succinate not normally present in cells, is a strong competitive inhibitor of succinate dehydrogenase, and its addition to mitochondria in the laboratory blocks the activity of the citric acid cycle.
Hydration of Fumarate to Malate
 Hydration of Fumarate to Malate
Hydration of Fumarate to MalateThe reversible hydration of fumarate to l-malate is catalyzed by fumarase (formally, fumarate hydratase). The transition state in this reaction is a carbanion:
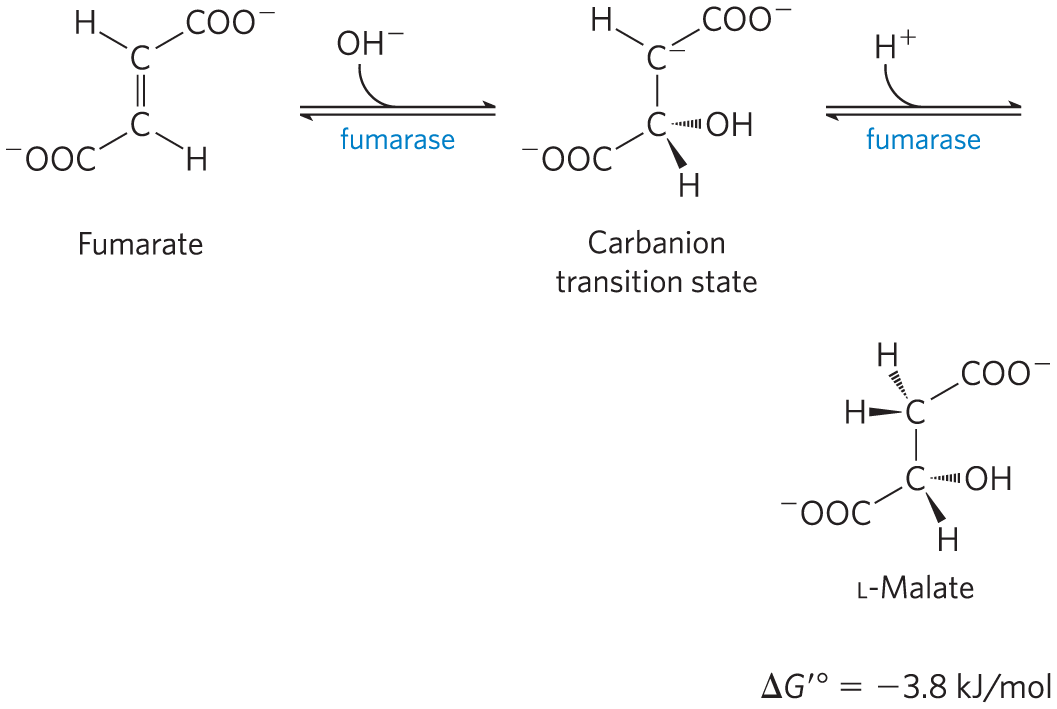
This enzyme is highly stereospecific; it catalyzes hydration of the trans double bond of fumarate but not the cis double bond of maleate. In the reverse direction (from l-malate to fumarate), fumarase is equally stereospecific: d-malate is not a substrate.

Oxidation of Malate to Oxaloacetate
In the last reaction of the citric acid cycle, l-malate dehydrogenase catalyzes the oxidation of l-malate to oxaloacetate, coupled to the reduction of to NADH:
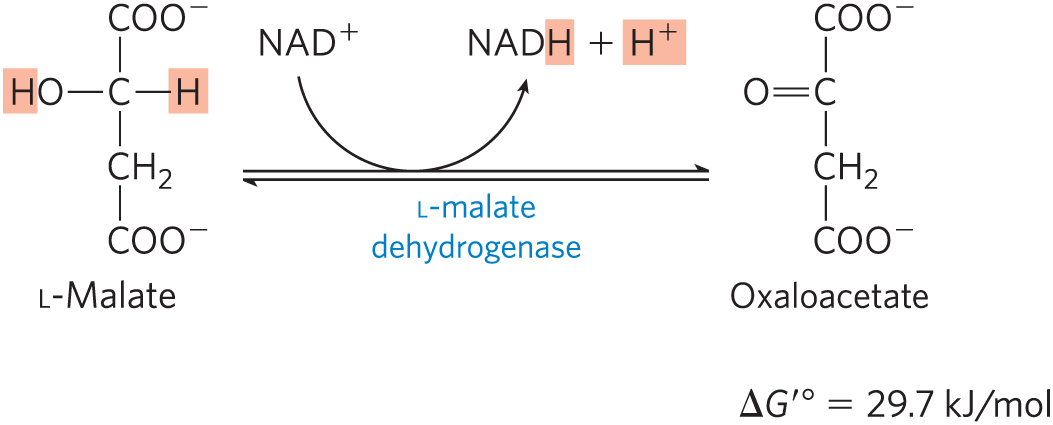
The equilibrium of this reaction lies far to the left under standard thermodynamic conditions, but in intact cells oxaloacetate is continually removed by the highly exergonic citrate synthase reaction (step of Fig. 16-7). This keeps the concentration of oxaloacetate in the cell extremely low , pulling the malate dehydrogenase reaction toward the formation of oxaloacetate.
Although the individual reactions of the citric acid cycle were initially worked out in vitro, using minced muscle tissue, the pathway and its regulation have also been studied extensively in vivo. By using precursors isotopically labeled with , researchers have traced the fate of individual carbon atoms through the citric acid cycle. Some of the earliest experiments with produced an unexpected result, however, which aroused considerable controversy about the pathway and mechanism of the citric acid cycle. In fact, these experiments at first seemed to show that citrate was not the first tricarboxylic acid to be formed. Box 16-2 gives some details of this episode in the history of citric acid cycle research. Today biochemists use -labeled precursors and whole-tissue NMR spectroscopy to monitor metabolic flux through the cycle in living tissue. Because the NMR signal is unique to the compound containing the , precursor carbons can be traced into each cycle intermediate and into compounds derived from the intermediates. This technique makes it possible to study regulation of the citric acid cycle and its interconnections with other metabolic pathways.
The Energy of Oxidations in the Cycle Is Efficiently Conserved
We have now covered one complete turn of the citric acid cycle (Fig. 16-14). A two-carbon acetyl group entered the cycle by combining with oxaloacetate. Two carbon atoms emerged from the cycle as from the oxidation of isocitrate and α-ketoglutarate. The energy released by these oxidations was conserved in the reduction of three and one FAD and the production of one ATP or GTP. At the end of the cycle a molecule of oxaloacetate was regenerated. Note that the two carbon atoms appearing as are not the same two carbons that entered in the form of the acetyl group; additional turns around the cycle are required to release these carbons as (Fig. 16-7).
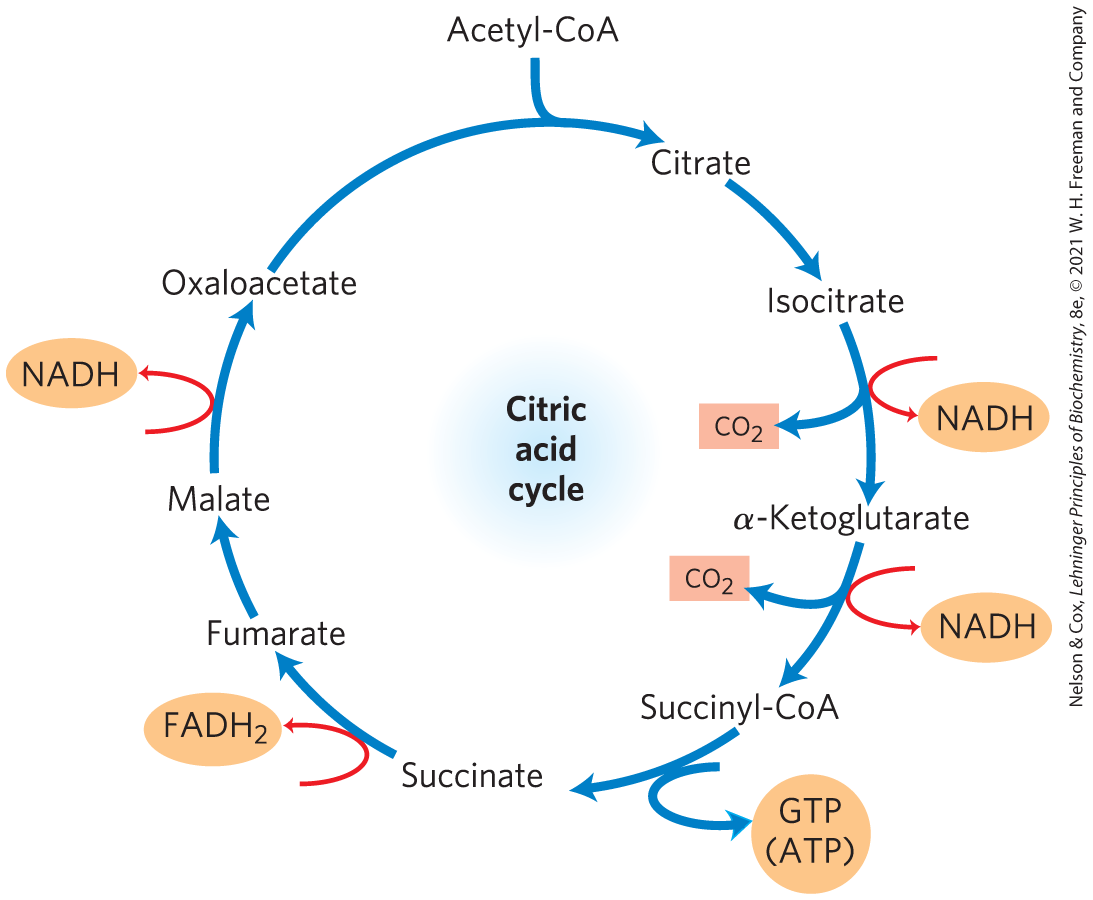
FIGURE 16-14 Products of one turn of the citric acid cycle. At each turn of the cycle, three NADH, one , one GTP (or ATP), and two are released in oxidative decarboxylation reactions. Here and in several of the following figures, all cycle reactions are shown as proceeding in one direction only, but keep in mind that most of the reactions are reversible.
Although the citric acid cycle directly generates only one ATP per turn (in the conversion of succinyl-CoA to succinate), the four oxidation steps in the cycle provide a large flow of electrons into the respiratory chain via NADH and and thus lead to formation of almost 10 times more ATP during oxidative phosphorylation.
We saw in Chapter 14 that the production of two molecules of pyruvate from one molecule of glucose in glycolysis yields 2 ATP and 2 NADH. In oxidative phosphorylation (Chapter 19), passage of two electrons from NADH to drives the formation of about 2.5 ATP, and passage of two electrons from to yields about 1.5 ATP. This stoichiometry allows us to calculate the overall yield of ATP from the complete oxidation of glucose. When both pyruvate molecules are oxidized to 6 via the pyruvate dehydrogenase complex and the citric acid cycle, and the electrons are transferred to via oxidative phosphorylation, as many as 32 ATP are obtained per glucose (Table 16-1). In round numbers, this represents the conservation of , or 34% of the theoretical maximum of about 2,840 kJ/mol available from the complete oxidation of glucose. These calculations employ the standard free-energy changes; when corrected for the actual free energy required to form ATP within cells (see Worked Example 13-2, p. 480), the calculated efficiency of the process is closer to 65%. When cells under anaerobic conditions depend on glycolysis for ATP, one glucose yields just 2 ATP. Aerobic metabolism is far more effective in capturing the energy in glucose as a fuel.
| Reaction | Number of ATP or reduced coenzyme directly formed | Number of ATP ultimately formeda |
|---|---|---|
2 NADH |
3 or 5b |
|
2 ATP |
2 |
|
2 ATP |
2 |
|
2 NADH |
5 |
|
2 NADH |
5 |
|
2 NADH |
5 |
|
2 ATP (or 2 GTP) |
2 |
|
3 |
||
2 NADH |
5 |
|
Total |
30–32 |
|
|
aThis is calculated as 2.5 ATP per NADH and 1.5 ATP per . A negative value indicates consumption. bThis number is either 3 or 5, depending on the mechanism used to shuttle NADH equivalents from the cytosol to the mitochondrial matrix; see Figures 19-31 and 19-32. |
||
SUMMARY 16.2 Reactions of the Citric Acid Cycle
- The citric acid cycle (Krebs cycle, TCA cycle) is a nearly universal central catabolic pathway in which compounds derived from the breakdown of carbohydrates, fats, and proteins are oxidized to , with most of the energy of oxidation temporarily held in the electron carriers and NADH. During aerobic metabolism, these electrons are transferred to and the energy of electron flow is conserved as ATP.
- Acetyl-CoA enters the citric acid cycle as citrate synthase catalyzes its condensation with oxaloacetate to form citrate. In seven sequential reactions, including two decarboxylations, the citric acid cycle converts citrate to oxaloacetate and releases two . The pathway is cyclic in that the intermediates of the cycle are not used up; for each oxaloacetate consumed in the path, one is produced.
- For each acetyl-CoA oxidized by the citric acid cycle, the energy gain consists of three molecules of NADH, one , and one nucleoside triphosphate (either ATP or GTP).
 The citric acid cycle (Krebs cycle, TCA cycle) is a nearly universal central catabolic pathway in which compounds derived from the breakdown of carbohydrates, fats, and proteins are oxidized to , with most of the energy of oxidation temporarily held in the electron carriers and NADH. During aerobic metabolism, these electrons are transferred to and the energy of electron flow is conserved as ATP.
The citric acid cycle (Krebs cycle, TCA cycle) is a nearly universal central catabolic pathway in which compounds derived from the breakdown of carbohydrates, fats, and proteins are oxidized to , with most of the energy of oxidation temporarily held in the electron carriers and NADH. During aerobic metabolism, these electrons are transferred to and the energy of electron flow is conserved as ATP.