In glycolysis (from the Greek glykys, “sweet” or “sugar,” and lysis, “splitting”), a molecule of glucose is degraded in a series of enzyme-catalyzed reactions to yield two molecules of the three-carbon compound pyruvate. During the sequential reactions of glycolysis, some of the free energy released from glucose is conserved in the form of ATP and NADH. Glycolysis was the first metabolic pathway to be elucidated and is probably the best understood. From Eduard Buchner’s discovery in 1897 of fermentation in cell-free extracts of yeast until the elucidation of the whole pathway in yeast and in muscle in the 1930s, the reactions of glycolysis were a major focus of biochemical research. These discoveries showed that the reactions of life could be explained chemically, without reliance on a mystical life force. This philosophical shift led physiologist Jacques Loeb to observe in 1906, “The history of this problem is instructive, as it warns us against considering problems as beyond our reach because they have not yet found their solution.”1
The development of methods of enzyme purification, the discovery and recognition of the importance of coenzymes such as NAD, and the discovery of the pivotal metabolic role of ATP and other phosphorylated compounds all came out of studies of glycolysis. The glycolytic enzymes of many species have long since been purified and thoroughly studied.
Glycolysis is an almost universal central pathway of glucose catabolism, the pathway with the largest flux of carbon in most cells. The glycolytic breakdown of glucose is the sole source of metabolic energy in some mammalian tissues and cell types (erythrocytes, renal medulla, brain, and sperm, for example). Some plant tissues that are modified to store starch (such as potato tubers) and some aquatic plants (watercress, for example) derive most of their energy from glycolysis; many anaerobic microorganisms are entirely dependent on glycolysis.
In the course of evolution, the chemistry of the reactions of glycolysis has been completely conserved. Genome sequencing and structural studies have shown that the glycolytic enzymes of vertebrates are closely similar in amino acid sequence and three-dimensional structure to their homologs in yeast and spinach. Although some archaea and parasitic microorganisms lack one or more of the enzymes of glycolysis, they retain the core of the pathway. The glycolytic pathway, of central importance in itself, is governed by thermodynamic principles and regulatory mechanisms that are common to all pathways of cell metabolism. It serves as a model of principles we will revisit throughout Part II of this book.
An Overview: Glycolysis Has Two Phases
Before examining each step of the pathway in some detail, we take a look at glycolysis as a whole. As all sugar derivatives in glycolysis are the d isomers, we will usually omit the d designation except when emphasizing stereochemistry. The breakdown of the six-carbon glucose into two molecules of the three-carbon pyruvate occurs in 10 steps, the first 5 of which constitute the preparatory phase (Fig. 14-2a). In these reactions, glucose is first phosphorylated at the hydroxyl group on C-6 (step ). The glucose 6-phosphate thus formed is converted to fructose 6-phosphate (step ), which is again phosphorylated, this time at C-1, to yield fructose 1,6-bisphosphate (step ). For both phosphorylations, ATP is the phosphoryl group donor.
FIGURE 14-2 The two phases of glycolysis. For each molecule of glucose that passes through the preparatory phase (a), two molecules of glyceraldehyde 3-phosphate are formed; both pass through the payoff phase (b). Pyruvate is the end product of the second phase of glycolysis. For each glucose molecule, two ATP are consumed in the preparatory phase and four ATP are produced in the payoff phase, giving a net yield of two ATP and two NADH per molecule of glucose converted to pyruvate. The numbered reaction steps correspond to the numbered headings in the text discussion. Keep in mind that each phosphoryl group, represented here as , has two negative charges .
Fructose 1,6-bisphosphate is split to yield two different three-carbon molecules, dihydroxyacetone phosphate and glyceraldehyde 3-phosphate (step ); this is the “lysis” step that gives the pathway its name. The dihydroxyacetone phosphate is isomerized to form a second molecule of glyceraldehyde 3-phosphate (step ), ending the first phase of glycolysis. Note that two molecules of ATP are invested before the cleavage of glucose into two three-carbon pieces; there will be a good return on this investment. To summarize: in the preparatory phase of glycolysis the energy of ATP is invested, raising the free-energy content of the intermediates, and the carbon chains of all the metabolized hexoses are converted to a common product, glyceraldehyde 3-phosphate.
The energy gain comes in the payoff phase of glycolysis (Fig. 14-2b). Each molecule of glyceraldehyde 3-phosphate is oxidized and phosphorylated by inorganic phosphate (not by ATP) to form 1,3-bisphosphoglycerate (step ). Energy is then released as the two molecules of 1,3-bisphosphoglycerate are converted to two molecules of pyruvate (steps through ). Much of this energy is conserved by the coupled phosphorylation of four molecules of ADP to ATP. The net yield is two molecules of ATP per molecule of glucose used, because two molecules of ATP were invested in the preparatory phase. Energy is also conserved in the payoff phase in the formation of two molecules of the electron carrier NADH per molecule of glucose.
In the sequential reactions of glycolysis, three types of chemical transformations are particularly noteworthy: (1) degradation of the carbon skeleton of glucose to yield pyruvate; (2) phosphorylation of ADP to ATP by compounds with high phosphoryl group transfer potential, formed during glycolysis; and (3) transfer of a hydride ion to , forming NADH. The overall chemical logic of the pathway is described in Figure 14-3.
FIGURE 14-3 The chemical logic of the glycolytic pathway. In this simplified version of the pathway, each molecule is shown in a linear form, with carbon and hydrogen atoms not depicted, in order to highlight chemical transformations. Remember that glucose and fructose are present mostly in their cyclized forms in solution, although they are transiently present in linear form at the active sites of some of the enzymes in this pathway.
The preparatory phase, steps to , converts the six-carbon glucose into two three-carbon units, each of them phosphorylated. Oxidation of the three-carbon units is initiated in the payoff phase, steps to . To produce pyruvate, the chemical steps must occur in the order shown.
ATP and NADH Formation Coupled to Glycolysis
During glycolysis some of the energy of the glucose molecule is conserved in ATP, while much remains in the product, pyruvate. The overall equation for glycolysis is
(14-1)
For each molecule of glucose degraded to pyruvate, two molecules of ATP are generated from ADP and , and two molecules of NADH are produced by the reduction of . The reduction of (see Fig. 13-24) proceeds by the enzymatic transfer of a hydride ion from the aldehyde group of glyceraldehyde 3-phosphate to the nicotinamide ring of , yielding the reduced coenzyme NADH. The other hydrogen atom of the substrate molecule is released to the solution as .
We can now resolve the equation of glycolysis into two processes — the conversion of glucose to pyruvate, which is exergonic:
(14-2)
and the formation of ATP from ADP and , which is endergonic:
(14-3)
The sum of Equations 14-2 and 14-3 gives the overall standard free-energy change of glycolysis, :
Under standard conditions, and under the (nonstandard) conditions that prevail in a cell, glycolysis is an essentially irreversible process, driven to completion by a large net decrease in free energy.
Energy Remaining in Pyruvate
Glycolysis releases only a small fraction of the total available energy of the glucose molecule. The two molecules of pyruvate formed by glycolysis still contain most of the chemical potential energy of glucose, energy that can be extracted by oxidative reactions in the citric acid cycle (Chapter 16) and oxidative phosphorylation (Chapter 19) — aerobic processes. Under anaerobic conditions, pyruvate can be reduced to lactate or ethanol (Section 14.3). The oxidation of pyruvate is an important catabolic process, but pyruvate has anabolic fates as well. It can, for example, provide the carbon skeleton for the synthesis of the amino acid alanine or for the synthesis of fatty acids. We return to these anabolic reactions of pyruvate in later chapters.
Importance of Phosphorylated Intermediates
Each of the nine glycolytic intermediates between glucose and pyruvate is phosphorylated (Fig. 14-2). The phosphoryl groups have three functions.
Because the plasma membrane lacks transporters for phosphorylated sugars, the phosphorylated glycolytic intermediates cannot leave the cell. After the initial phosphorylation, no further energy is necessary to retain phosphorylated intermediates in the cell, despite the large difference in their intracellular and extracellular concentrations.
Phosphoryl groups are essential components in the enzymatic conservation of metabolic energy. Energy made available with the transfer of phosphoryl groups from compounds with phosphoanhydride bonds (such as those in ATP) is partially conserved in the formation of phosphate esters such as glucose 6-phosphate. Compounds with higher group transfer potential than ATP, which are formed in glycolysis (1,3-bisphosphoglycerate and phosphoenolpyruvate), donate phosphoryl groups to ADP to form ATP.
Binding energy resulting from the binding of phosphate groups to the active sites of enzymes lowers the activation energy and increases the specificity of the enzymatic reactions (Chapter 6). The phosphate groups of ADP, ATP, and the glycolytic intermediates form complexes with , and the substrate binding sites of many glycolytic enzymes are specific for these complexes. Most glycolytic enzymes require for activity.
The Preparatory Phase of Glycolysis Requires ATP
In the preparatory phase of glycolysis, two molecules of ATP are invested and the hexose chain is cleaved into two triose phosphates. The realization that phosphorylated hexoses were intermediates in glycolysis came slowly and serendipitously. In 1906, Arthur Harden and William Young tested their hypothesis that inhibitors of proteolytic enzymes would stabilize the glucose-fermenting enzymes in yeast extract. They added blood serum (known to contain inhibitors of proteolytic enzymes) to yeast extracts and observed the predicted stimulation of glucose metabolism. However, in a control experiment intended to show that boiling the serum destroyed the stimulatory activity, they discovered that boiled serum was just as effective at stimulating glycolysis! Careful examination and testing of the contents of the boiled serum revealed that inorganic phosphate was responsible for the stimulation. Harden and Young soon discovered that glucose added to their yeast extract was converted to a hexose bisphosphate (the “Harden-Young ester,” eventually identified as fructose 1,6-bisphosphate). This was the beginning of a long series of investigations on the role of organic esters and anhydrides of phosphate in biochemistry, which has led to our current understanding of the central role of phosphoryl group transfer in biology.
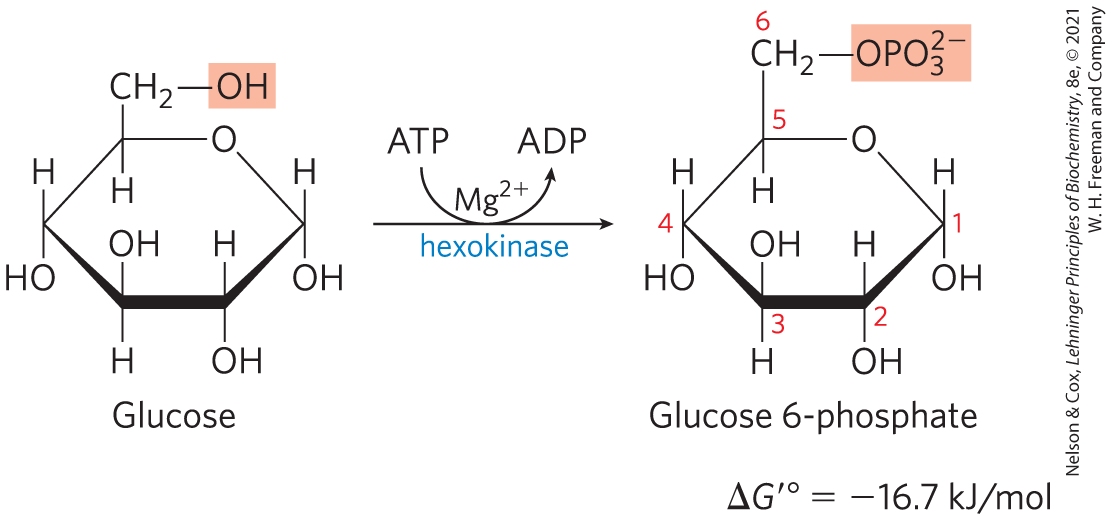
Phosphorylation of Glucose
In the first step of glycolysis (Fig. 14-2), glucose is activated for subsequent reactions by its phosphorylation at C-6 to yield glucose 6-phosphate, with ATP as the phosphoryl donor:
This reaction, which is irreversible under intracellular conditions, is catalyzed by hexokinase. Recall that kinases are enzymes that catalyze the transfer of the terminal phosphoryl group from ATP to an acceptor nucleophile (see Fig. 13-8c). Kinases are a subclass of transferases (see Table 6-3). The acceptor in the case of hexokinase is a hexose, normally glucose, although hexokinase also catalyzes the phosphorylation of other common hexoses, such as fructose and mannose, in some tissues.
Hexokinase, like many other kinases, requires for its activity, because the true substrate of the enzyme is not but the complex (see Fig. 13-12). shields the negative charges of the phosphoryl groups in ATP, making the terminal phosphorus atom an easier target for nucleophilic attack by an of glucose. Hexokinase undergoes a profound change in shape, an induced fit, when it binds glucose; two domains of the protein move about 8 Å closer to each other when ATP binds (see Fig. 6-30). This movement brings bound ATP closer to a molecule of glucose also bound to the enzyme and blocks the access of water (from the solvent), which might otherwise enter the active site and attack (hydrolyze) the phosphoanhydride bonds of ATP. Like the other nine enzymes of glycolysis, hexokinase is a soluble, cytosolic protein.
Hexokinase is present in nearly all organisms. The human genome encodes four different hexokinases (I to IV), all of which catalyze the same reaction, but differ in kinetics, regulation, and location. Two or more enzymes that catalyze the same reaction but are encoded by different genes are called isozymes (see Box 14-3). One of the isozymes present in hepatocytes, hexokinase IV (also called glucokinase), differs from other forms of hexokinase in kinetic and regulatory properties, with important physiological consequences that are described in Section 14.5.
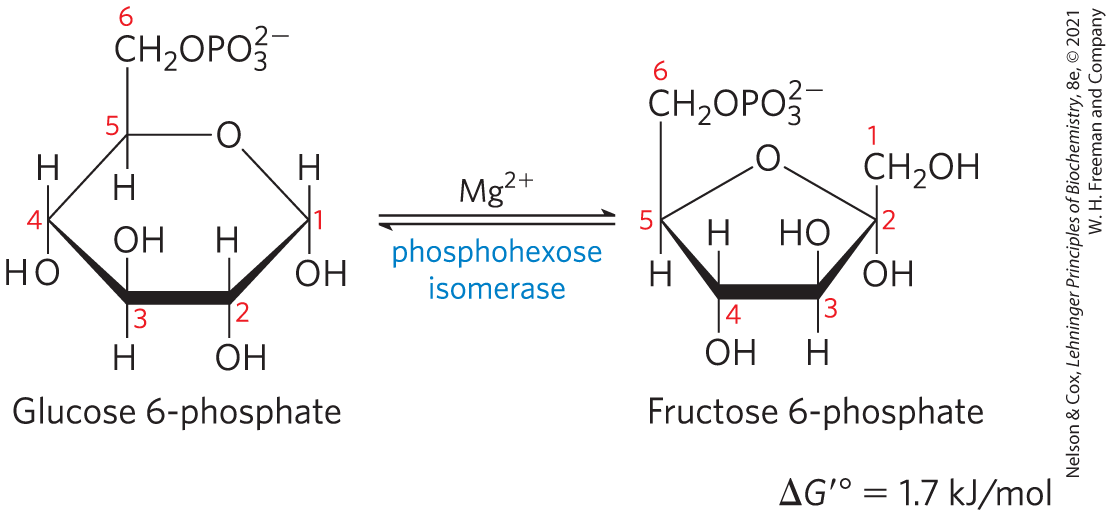
Conversion of Glucose 6-Phosphate to Fructose 6-Phosphate
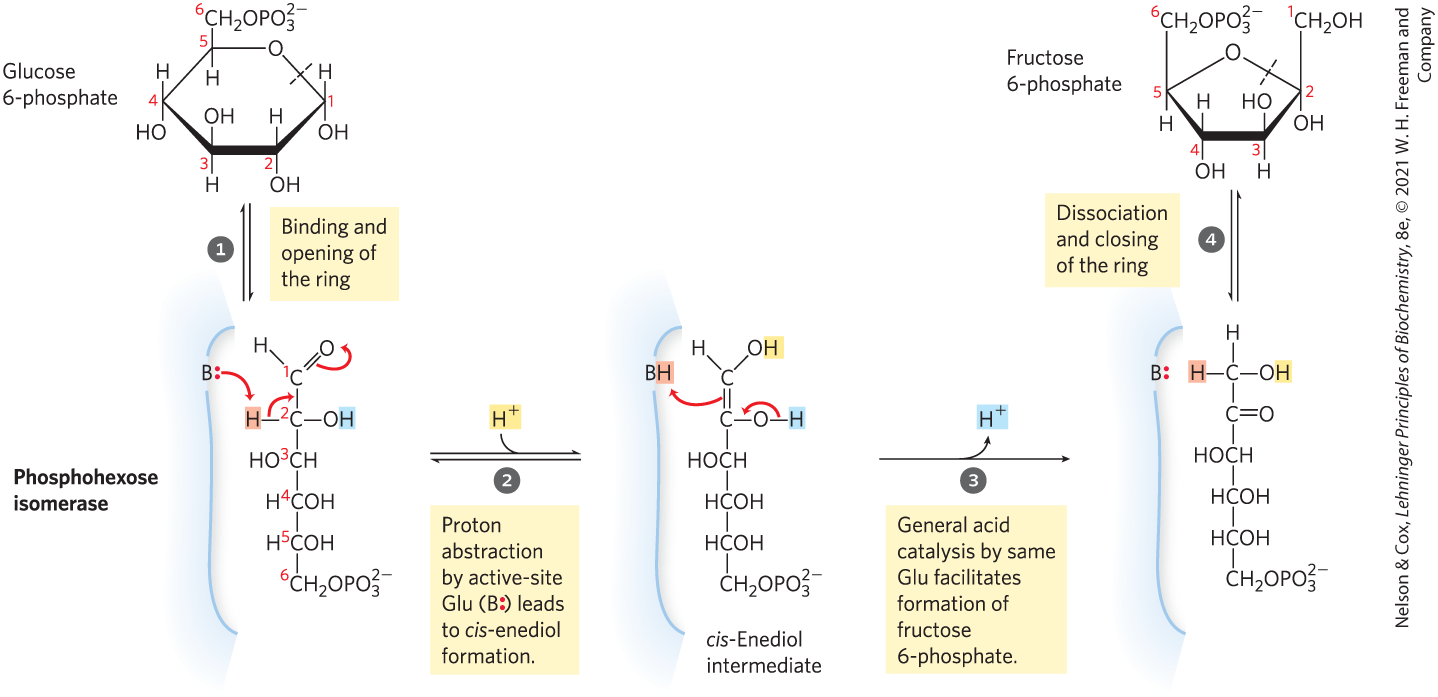
The enzyme phosphohexose isomerase (phosphoglucose isomerase) catalyzes the reversible isomerization of glucose 6-phosphate, an aldose, to fructose 6-phosphate, a ketose:
The mechanism for this reaction involves an enediol intermediate (Fig. 14-4). The reaction proceeds readily in either direction, as might be expected from the relatively small change in standard free energy.
MECHANISM FIGURE 14-4 The phosphohexose isomerase reaction. The ring opening and closing reactions (steps and ) are catalyzed by an active-site His residue, by mechanisms omitted here for simplicity. The proton (light red) initially at C-2 is made more easily abstractable by electron withdrawal by the adjacent carbonyl and nearby hydroxyl groups. After its transfer from C-2 to the active-site Glu residue (a weak acid), the proton is freely exchanged with the surrounding solution; that is, the proton abstracted from C-2 in step is not necessarily the same one that is added to C-1 in step .
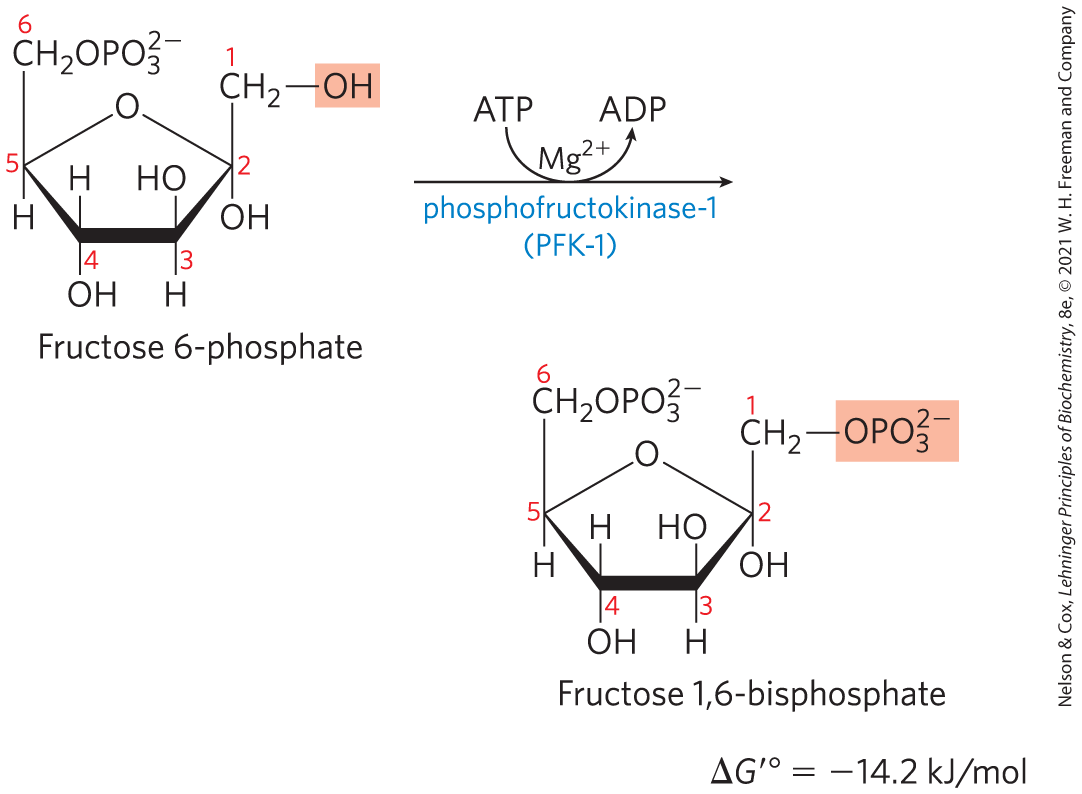
Phosphorylation of Fructose 6-Phosphate to Fructose 1,6-Bisphosphate
In the second of the two priming reactions of glycolysis, phosphofructokinase-1 (PFK-1) catalyzes the transfer of a phosphoryl group from ATP to fructose 6-phosphate to yield fructose 1,6-bisphosphate:
Key convention
Compounds that contain two phosphate or phosphoryl groups attached at different positions in the molecule are named bisphosphates (or bisphospho compounds); for example, fructose 1,6-bisphosphate and 1,3-bisphosphoglycerate. Compounds with two phosphates linked together as a pyrophosphoryl group are named diphosphates; for example, adenosine diphosphate (ADP). Similar rules apply for the naming of trisphosphates (such as inositol 1,4,5-trisphosphate; see p. 425) and triphosphates (such as adenosine triphosphate, ATP).
The enzyme that forms fructose 1,6-bisphosphate is called PFK-1 to distinguish it from a second enzyme (PFK-2) that catalyzes the formation of fructose 2,6-bisphosphate from fructose 6-phosphate in a separate pathway (the roles of PFK-2 and fructose 2,6-bisphosphate are discussed in Section 14.5). The PFK-1 reaction is essentially irreversible under cellular conditions, and it is the first “committed” step in the glycolytic pathway; glucose 6-phosphate and fructose 6-phosphate have other possible fates, but fructose 1,6-bisphosphate is targeted for glycolysis.
Phosphofructokinase-1 is subject to complex allosteric regulation; its activity is increased whenever the cell’s ATP supply is depleted or when the ATP breakdown products, ADP and AMP (particularly the latter), accumulate. The enzyme is inhibited whenever the cell has ample ATP and is well supplied by other fuels such as fatty acids. In some organisms, fructose 2,6-bisphosphate (not to be confused with the PFK-1 reaction product, fructose 1,6-bisphosphate) is a potent allosteric activator of PFK-1. Ribulose 5-phosphate, an intermediate in the pentose phosphate pathway discussed in Section 14.6, also activates phosphofructokinase indirectly. The multiple layers of regulation of this step in glycolysis are discussed in greater detail in Section 14.5.
Some bacteria and protists and perhaps all plants have a different phosphofructokinase (PP-PFK-1) that uses pyrophosphate , not ATP, as the phosphoryl group donor in the synthesis of fructose 1,6-bisphosphate:
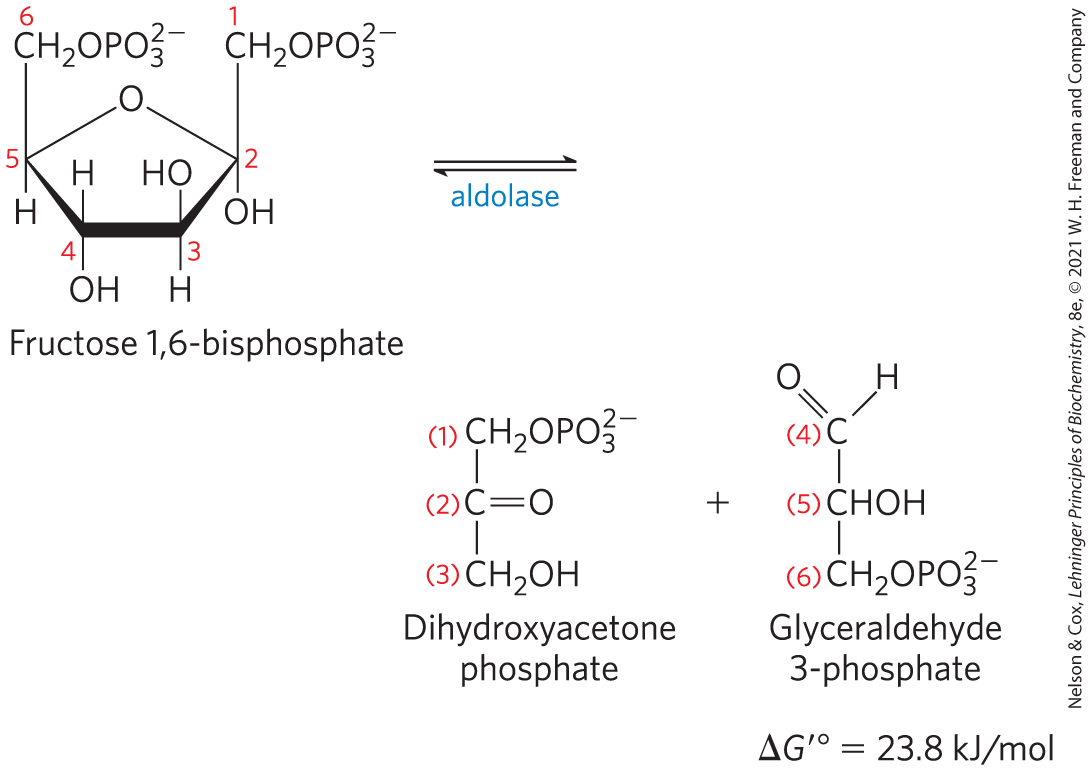
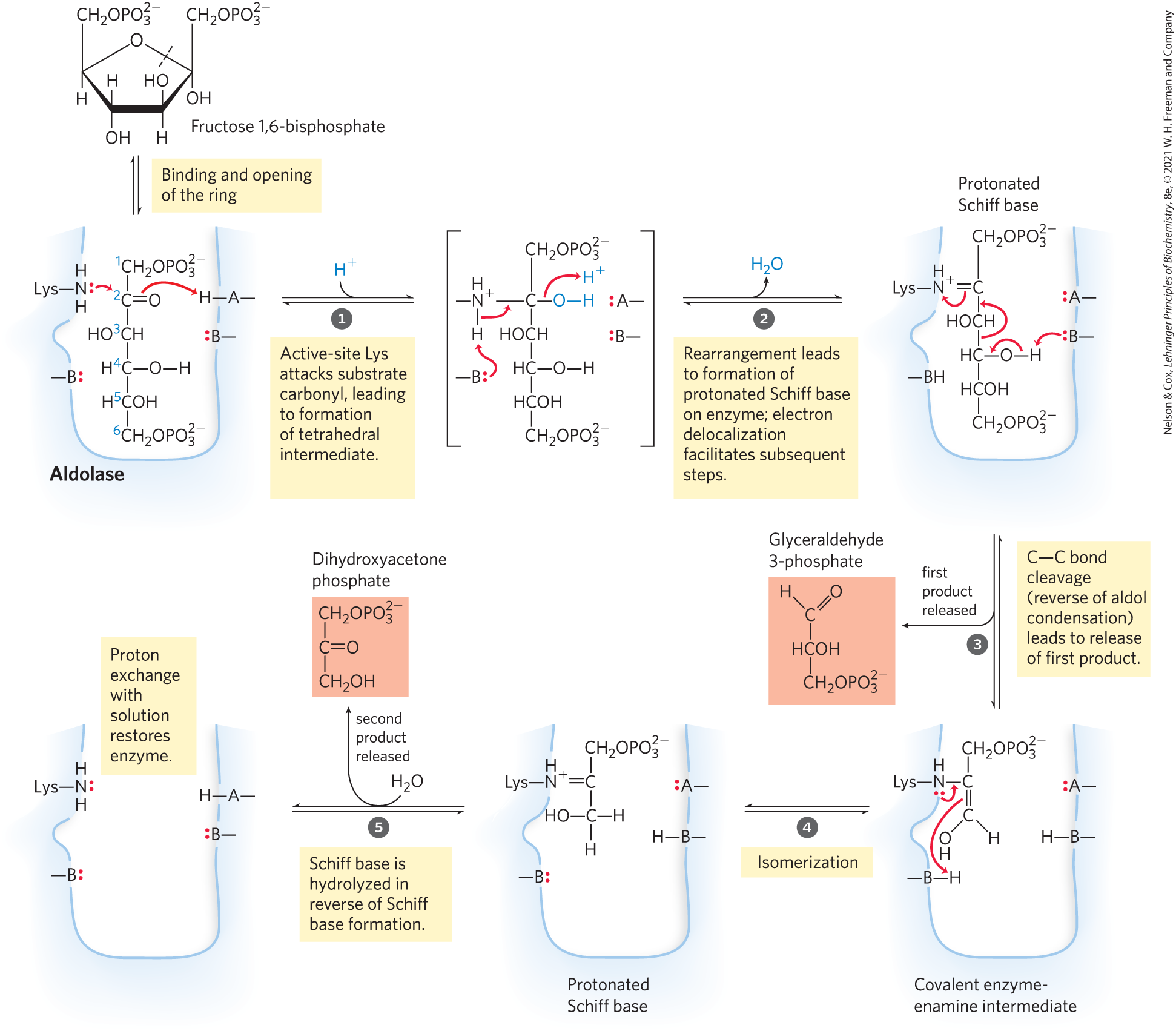
The enzyme fructose 1,6-bisphosphate aldolase, often called simply aldolase, catalyzes a reverse aldol condensation (Fig. 14-5; see Fig. 13-4). Fructose 1,6-bisphosphate is cleaved to yield two different triose phosphates, namely, glyceraldehyde 3-phosphate (an aldose) and dihydroxyacetone phosphate (a ketose):
MECHANISM FIGURE 14-5 The class I aldolase reaction. Note that cleavage between C-3 and C-4 depends on the presence of the carbonyl group at C-2, which is converted to an imine on the enzyme. A and B represent amino acid residues that serve as general acid (A) or base (B).
There are two classes of aldolases. Class I aldolases, found in animals and plants, use the mechanism shown in Figure 14-5. Class II enzymes, in fungi and bacteria, do not form the Schiff base intermediate. Instead, a zinc ion at the active site is coordinated with the carbonyl oxygen at C-2; the polarizes the carbonyl group and stabilizes the enolate intermediate created in the bond cleavage step (see Fig. 6-23).
Although the aldolase reaction has a strongly positive standard free-energy change in the direction of fructose 1,6-bisphosphate cleavage, at the lower concentrations of reactants present in cells the actual free-energy change is small and the aldolase reaction is readily reversible. We shall see later that aldolase acts in the reverse direction during the process of gluconeogenesis.
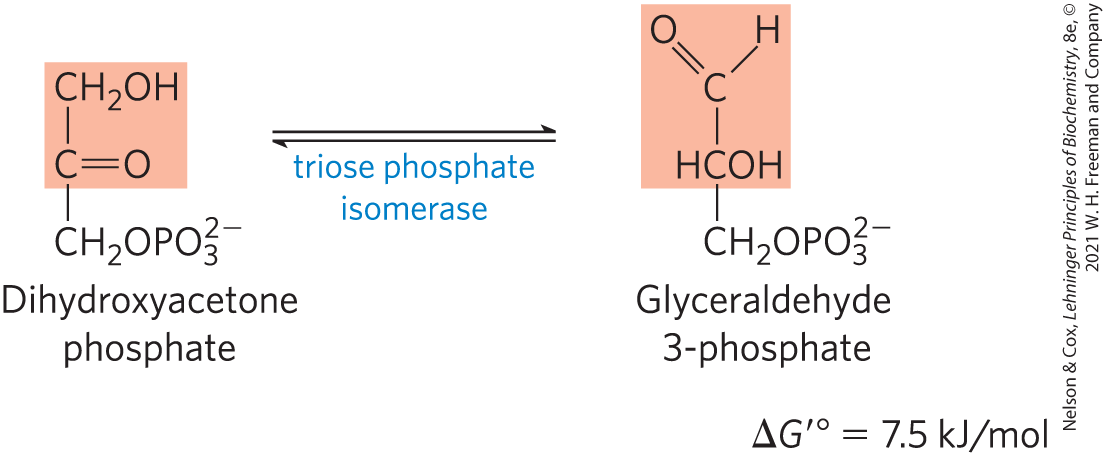
Interconversion of the Triose Phosphates
Only one of the two triose phosphates formed by aldolase, glyceraldehyde 3-phosphate, can be directly degraded in the subsequent steps of glycolysis. The other product, dihydroxyacetone phosphate, is immediately and reversibly converted to glyceraldehyde 3-phosphate by the fifth enzyme of the glycolytic sequence, triose phosphate isomerase:
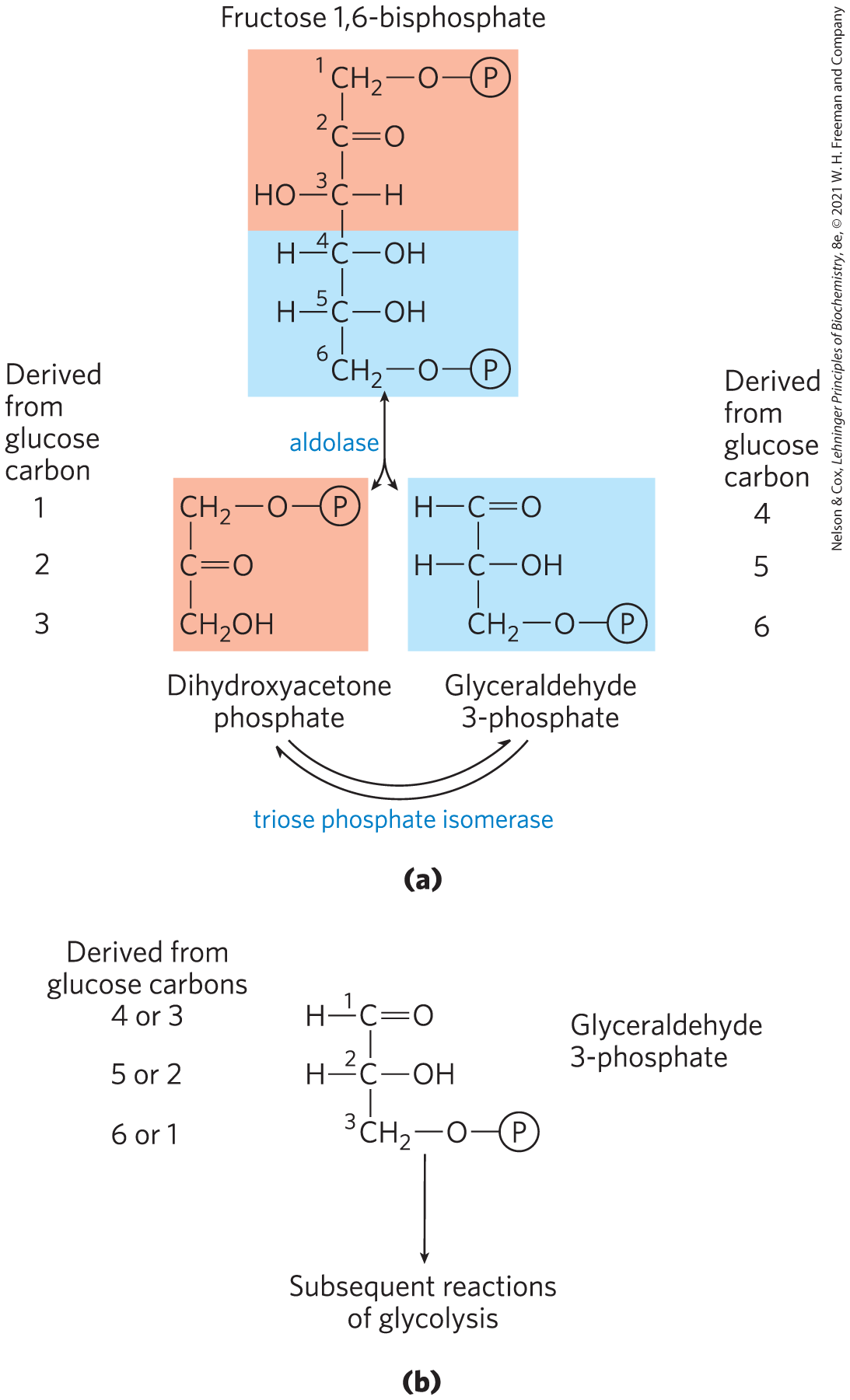
The reaction mechanism is similar to the reaction promoted by phosphohexose isomerase in step of glycolysis (Fig. 14-4). After the triose phosphate isomerase reaction, the carbon atoms derived from C-1, C-2, and C-3 of the starting glucose are chemically indistinguishable from C-6, C-5, and C-4, respectively (Fig. 14-6); both “halves” of glucose have yielded glyceraldehyde 3-phosphate.
FIGURE 14-6 Fate of the glucose carbons in the formation of glyceraldehyde 3-phosphate. (a) The origin of the carbons in the two three-carbon products of the aldolase and triose phosphate isomerase reactions. The end product of the two reactions is glyceraldehyde 3-phosphate (two molecules). (b) Each carbon of glyceraldehyde 3-phosphate is derived from either of two specific carbons of glucose. Note that the numbering of the carbon atoms of glyceraldehyde 3-phosphate differs from that of the glucose from which it is derived. In glyceraldehyde 3-phosphate, the most complex functional group (the carbonyl) is specified as C-1. This numbering change is important for interpreting experiments with glucose in which a single carbon is labeled with a radioisotope. (See Problems 5 and 22 at the end of this chapter.)
This reaction completes the preparatory phase of glycolysis. The hexose molecule has been phosphorylated at C-1 and C-6 and then cleaved to form two molecules of glyceraldehyde 3-phosphate.
The Payoff Phase of Glycolysis Yields ATP and NADH
The payoff phase of glycolysis (Fig. 14-2b) includes the energy-conserving phosphorylation steps in which some of the chemical energy of the glucose molecule is conserved in the form of ATP and NADH. Remember that one molecule of glucose yields two molecules of glyceraldehyde 3-phosphate, and both halves of the glucose molecule follow the same pathway in the second phase of glycolysis. The conversion of two molecules of glyceraldehyde 3-phosphate to two molecules of pyruvate is accompanied by the formation of four molecules of ATP from ADP. However, the net yield of ATP per molecule of glucose degraded is only two, because two ATP were invested in the preparatory phase of glycolysis to phosphorylate the two ends of the hexose molecule.
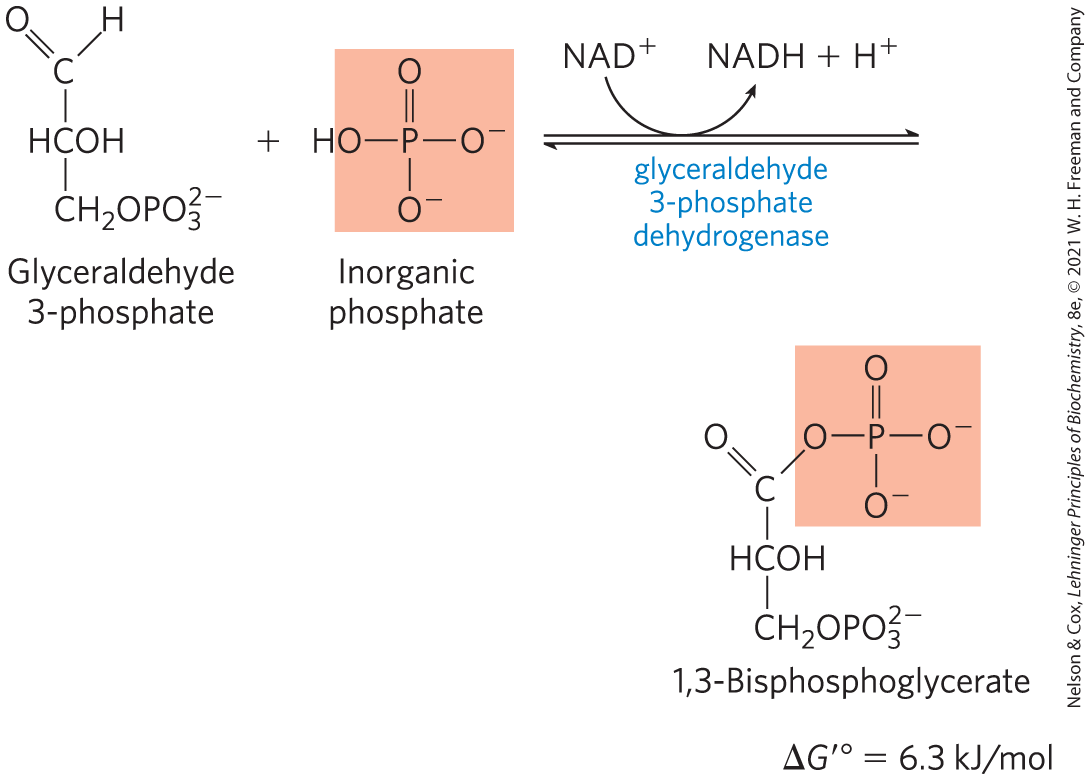
Oxidation of Glyceraldehyde 3-Phosphate to 1,3-Bisphosphoglycerate
The first step in the payoff phase is the oxidation of glyceraldehyde 3-phosphate to 1,3-bisphosphoglycerate, catalyzed by glyceraldehyde 3-phosphate dehydrogenase:
This is the first of the two energy-conserving reactions of glycolysis that eventually lead to the formation of ATP. The aldehyde group of glyceraldehyde 3-phosphate is oxidized, not to a free carboxyl group but to a carboxylic acid anhydride with phosphoric acid. This type of anhydride, called an acyl phosphate, has a very high standard free energy of hydrolysis (; see Table 13-6). Much of the free energy of oxidation of the aldehyde group of glyceraldehyde 3-phosphate is conserved by formation of the acyl phosphate group at C-1 of 1,3-bisphosphoglycerate.
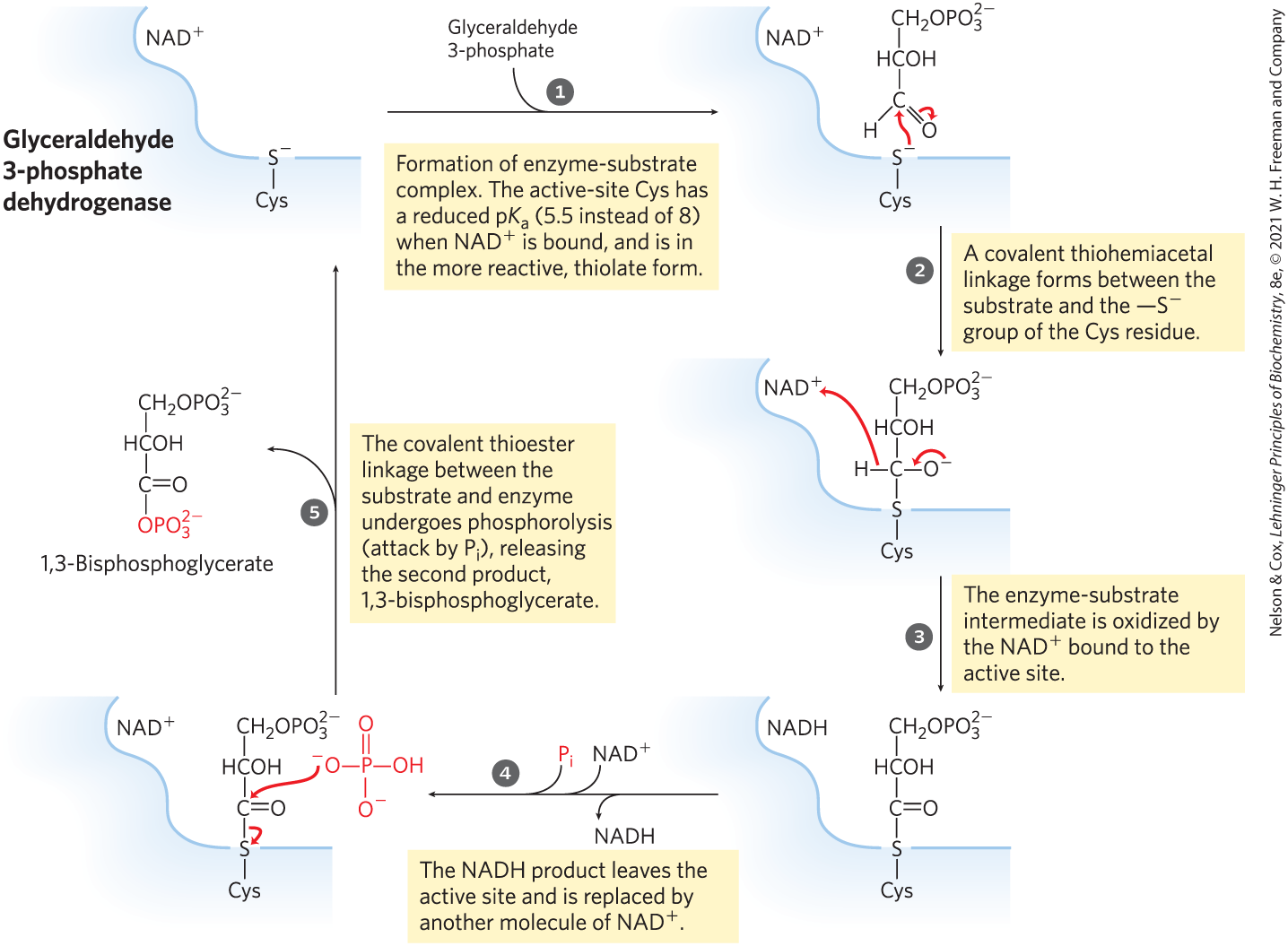
Glyceraldehyde 3-phosphate is covalently bound to the dehydrogenase during the reaction (Fig. 14-7). The aldehyde group of glyceraldehyde 3-phosphate reacts with the group of an essential Cys residue in the active site, in a reaction analogous to the formation of a hemiacetal (see Fig. 7-5), in this case producing a thiohemiacetal. Reaction of the essential Cys residue with a heavy metal such as irreversibly inhibits the enzyme.
MECHANISM FIGURE 14-7 The glyceraldehyde 3-phosphate dehydrogenase reaction.
The amount of in a cell is far smaller than the amount of glucose metabolized in a few minutes. Glycolysis would soon come to a halt if the NADH formed in this step of glycolysis were not continuously reoxidized and recycled. We return to a discussion of this recycling of later in the chapter.
Phosphoryl Transfer from 1,3-Bisphosphoglycerate to ADP
The enzyme phosphoglycerate kinase transfers the high-energy phosphoryl group from the carboxyl group of 1,3-bisphosphoglycerate to ADP, forming ATP and 3-phosphoglycerate:
Notice that phosphoglycerate kinase is named for the reverse reaction, in which it transfers a phosphoryl group from ATP to 3-phosphoglycerate. Like all enzymes, it catalyzes the reaction in both directions. This enzyme acts in the direction suggested by its name during gluconeogenesis (see Fig. 14-16) and during photosynthetic assimilation (see Fig. 20-26). In glycolysis, the reaction it catalyzes proceeds as shown above, in the direction of ATP synthesis.
Steps and of glycolysis together constitute an energy-coupling process in which 1,3-bisphosphoglycerate is the common intermediate; it is formed in the first reaction (which would be endergonic in isolation), and its acyl phosphate group is transferred to ADP in the second reaction (which is strongly exergonic). The sum of these two reactions is
Thus the overall reaction is exergonic.
Recall from Chapter 13 that the actual free-energy change, , is determined by the standard free-energy change, , and the mass-action ratio, Q, which is the ratio [products]/[reactants] (see Eqn 13-4). For step ,
Notice that is not included in Q. In biochemical calculations, is assumed to be a constant , and this constant is included in the definition of (p. 468).
When the mass-action ratio is less than 1.0, its natural logarithm has a negative sign. In the cytosol, where these reactions are taking place, the ratio is a small fraction, contributing to a low Q. Step , by consuming the product of step (1,3-bisphosphoglycerate), keeps [1,3-bisphosphoglycerate] relatively low in the steady state and thereby keeps Q for the overall energy-coupling process small. When Q is small, the contribution of ln Q can make strongly negative. This is simply another way of showing how the two reactions, steps and , are coupled through a common intermediate.
The outcome of these coupled reactions, both reversible under cellular conditions, is that the energy released on oxidation of an aldehyde to a carboxylate group is conserved by the coupled formation of ATP from ADP and . The formation of ATP by phosphoryl group transfer from a substrate such as 1,3-bisphosphoglycerate is referred to as a substrate-level phosphorylation, to distinguish this mechanism from respiration-linked phosphorylation. Substrate-level phosphorylations involve soluble enzymes and chemical intermediates (1,3-bisphosphoglycerate in this case). Respiration-linked phosphorylations, on the other hand, involve membrane-bound enzymes and transmembrane gradients of protons (Chapter 19).
Conversion of 3-Phosphoglycerate to 2-Phosphoglycerate
The enzyme phosphoglycerate mutase catalyzes a reversible shift of the phosphoryl group between C-2 and C-3 of glycerate; is essential for this reaction:
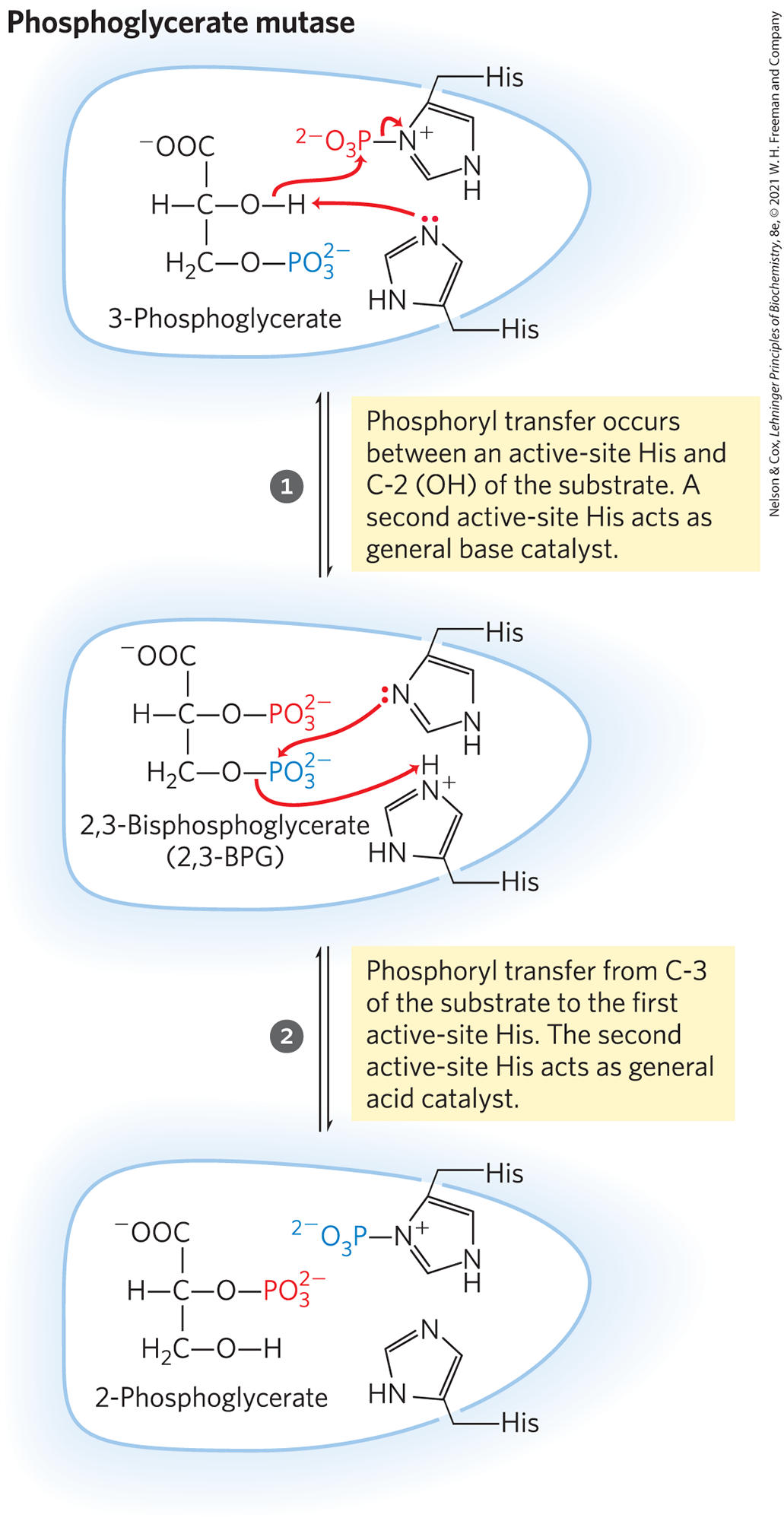
The reaction occurs in two steps (Fig. 14-8). A phosphoryl group initially attached to a His residue of the mutase is transferred to the hydroxyl group at C-2 of 3-phosphoglycerate, forming 2,3-bisphosphoglycerate (2,3-BPG). The phosphoryl group at C-3 of 2,3-BPG is then transferred to the same His residue, producing 2-phosphoglycerate and regenerating the phosphorylated enzyme. Phosphoglycerate mutase is initially phosphorylated by phosphoryl transfer from 2,3-BPG, which is required in small quantities to initiate the catalytic cycle and is continuously regenerated by that cycle.
MECHANISM FIGURE 14-8 The phosphoglycerate mutase reaction.
Dehydration of 2-Phosphoglycerate to Phosphoenolpyruvate
In the second glycolytic reaction that generates a compound with high phosphoryl group transfer potential (the first was step ), enolase promotes reversible removal of a molecule of water from 2-phosphoglycerate to yield phosphoenolpyruvate (PEP):
The mechanism of the enolase reaction involves an enolic intermediate stabilized by (see Fig. 6-31). The reaction converts a compound with a relatively low phosphoryl group transfer potential for hydrolysis of 2-phosphoglycerate is to one with high phosphoryl group transfer potential for PEP hydrolysis is (see Fig. 13-13).
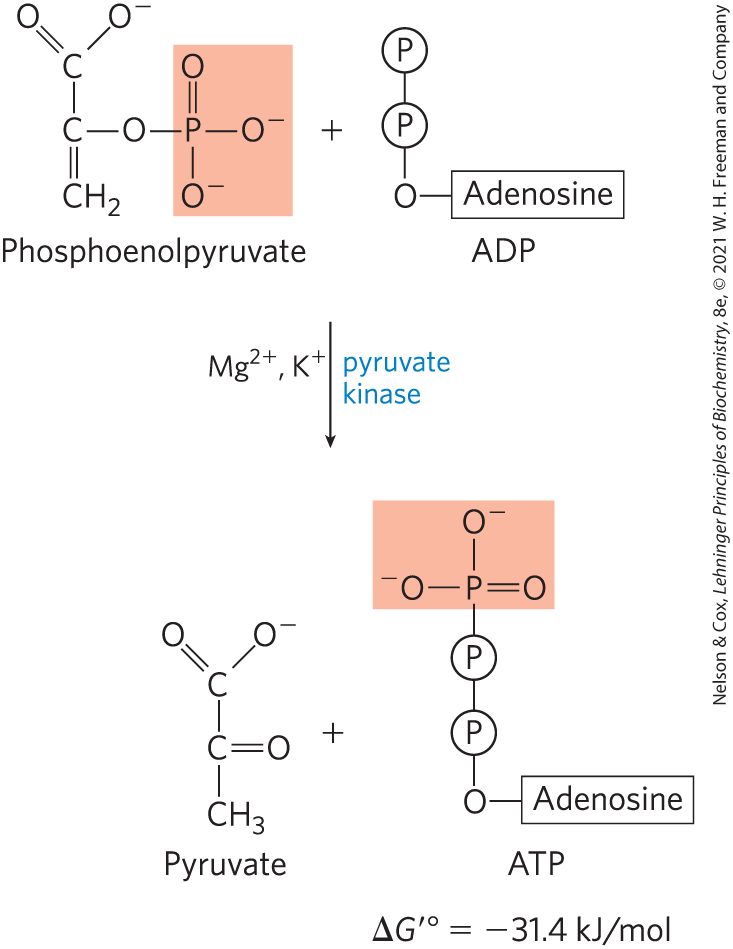
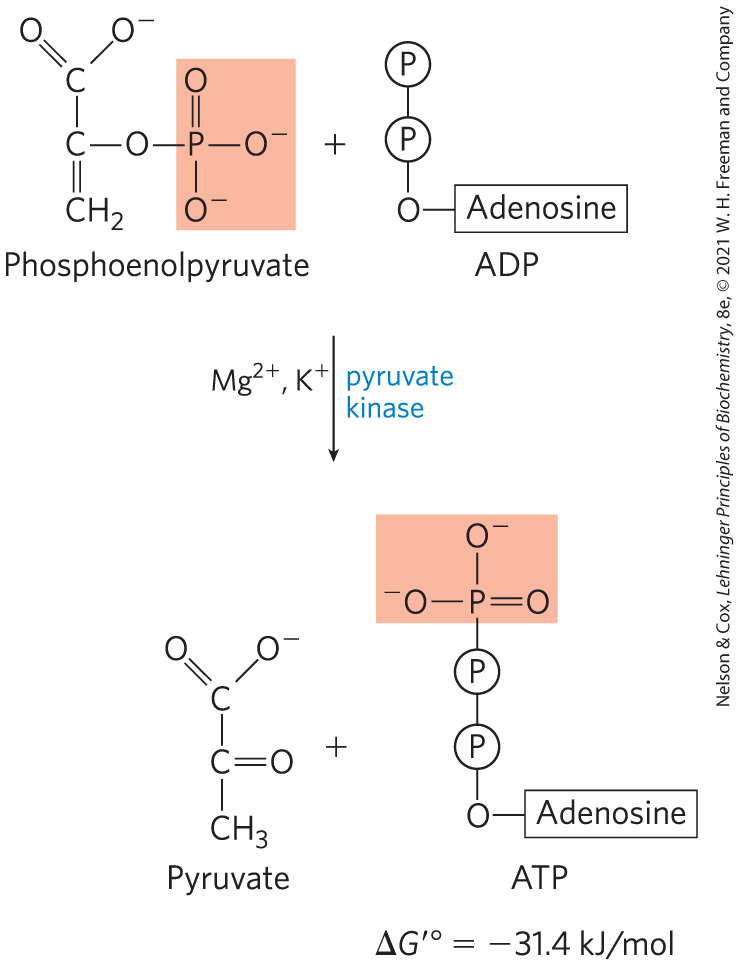
Transfer of the Phosphoryl Group from Phosphoenolpyruvate to ADP
The last step in glycolysis is the transfer of the phosphoryl group from phosphoenolpyruvate to ADP, catalyzed by pyruvate kinase, which requires and either or :
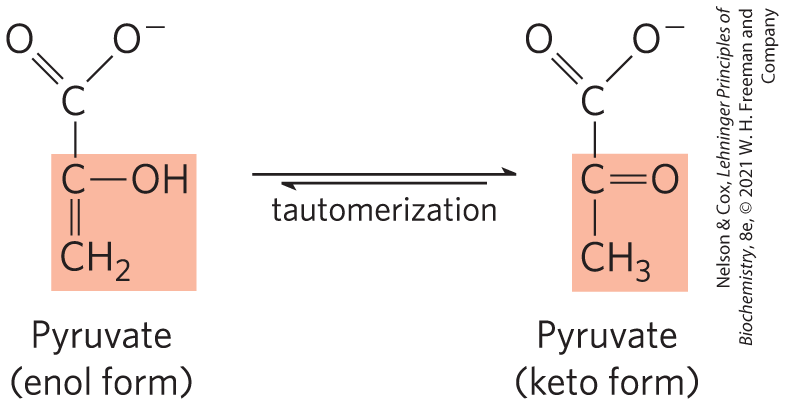
In this substrate-level phosphorylation, the product pyruvate first appears in its enol form, then tautomerizes nonenzymatically to its keto form, which predominates at pH 7:
The overall reaction has a large, negative standard free-energy change, due in large part to the spontaneous conversion of the enol form of pyruvate to the keto form (see Fig. 13-13). About half of the energy released by PEP hydrolysis is conserved in the formation of the phosphoanhydride bond of ATP , and the rest constitutes a large driving force pushing the reaction toward ATP synthesis. We discuss the regulation of pyruvate kinase in Section 14.5.
The Overall Balance Sheet Shows a Net Gain of Two ATP and Two NADH Per Glucose
We can now construct a balance sheet for glycolysis to account for (1) the fate of the carbon skeleton of glucose, (2) the input of and ADP and output of ATP, and (3) the pathway of electrons in the oxidation-reduction reactions. The left side of the following equation shows all the inputs of ATP, , ADP, and (consult Fig. 14-2), and the right side shows all the outputs (keep in mind that each molecule of glucose yields two molecules of pyruvate):
Canceling out common terms on both sides of the equation gives the overall equation for glycolysis:
In the overall glycolytic process, one molecule of glucose is converted to two molecules of pyruvate (the pathway of carbon). Two molecules of ADP and two of are converted to two molecules of ATP (the pathway of phosphoryl groups). Four electrons, as two hydride ions, are transferred from two molecules of glyceraldehyde 3-phosphate to two of (the pathway of electrons).
SUMMARY 14.1 Glycolysis
Glycolysis is a near-universal pathway by which a glucose molecule is oxidized, in two phases, to two molecules of pyruvate, with energy conserved as ATP and NADH. Ten cytosolic enzymes act sequentially in glycolysis. The overall reaction converts glucose to two molecules of pyruvate, and energy is conserved in the synthesis of two molecules of ATP and two molecules of NADH.
In the preparatory phase of glycolysis, two molecules of ATP are invested to activate glucose to fructose 1,6-bisphosphate. The bond between C-3 and C-4 is then broken to yield two molecules of triose phosphate.
In the payoff phase, each of the two molecules of glyceraldehyde 3-phosphate derived from glucose undergoes oxidation at C-1; some of the energy of this oxidation reaction is conserved in the form of one NADH and two ATP per triose phosphate oxidized.
Subtracting the two ATP spent in the preparatory phase, the net equation for the overall process is
 In
In  ). The glucose 6-phosphate thus formed is converted to fructose 6-phosphate (step
). The glucose 6-phosphate thus formed is converted to fructose 6-phosphate (step  ), which is again phosphorylated, this time at C-1, to yield fructose 1,6-bisphosphate (step
), which is again phosphorylated, this time at C-1, to yield fructose 1,6-bisphosphate (step  ). For both phosphorylations, ATP is the phosphoryl group donor.
). For both phosphorylations, ATP is the phosphoryl group donor.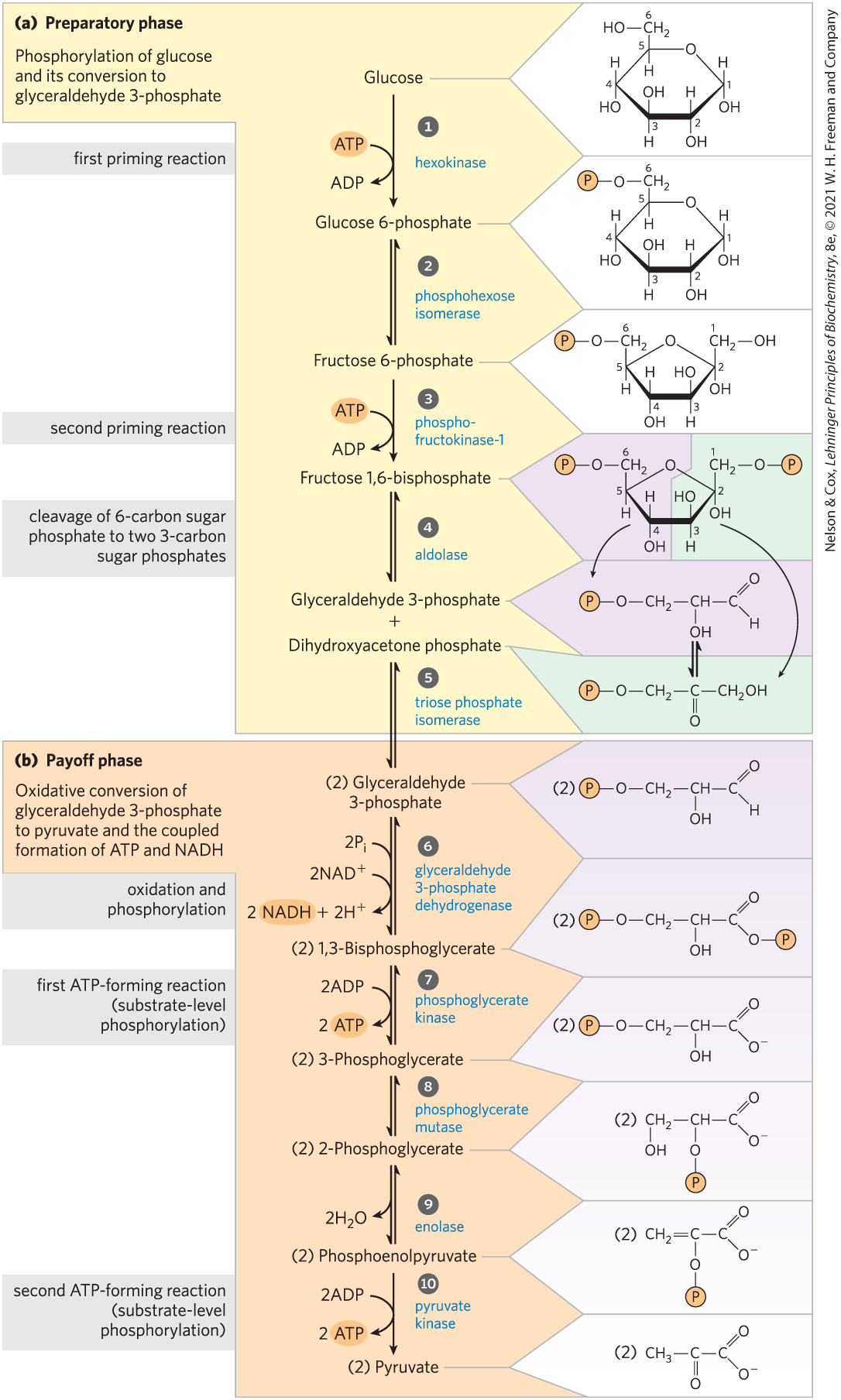
 , has two negative charges
, has two negative charges  ); this is the “lysis” step that gives the pathway its name. The dihydroxyacetone phosphate is isomerized to form a second molecule of glyceraldehyde 3-phosphate (step
); this is the “lysis” step that gives the pathway its name. The dihydroxyacetone phosphate is isomerized to form a second molecule of glyceraldehyde 3-phosphate (step  ), ending the first phase of glycolysis. Note that two molecules of ATP are invested before the cleavage of glucose into two three-carbon pieces; there will be a good return on this investment. To summarize: in the preparatory phase of glycolysis the energy of ATP is invested, raising the free-energy content of the intermediates, and the carbon chains of all the metabolized hexoses are converted to a common product, glyceraldehyde 3-phosphate.
), ending the first phase of glycolysis. Note that two molecules of ATP are invested before the cleavage of glucose into two three-carbon pieces; there will be a good return on this investment. To summarize: in the preparatory phase of glycolysis the energy of ATP is invested, raising the free-energy content of the intermediates, and the carbon chains of all the metabolized hexoses are converted to a common product, glyceraldehyde 3-phosphate. ). Energy is then released as the two molecules of 1,3-bisphosphoglycerate are converted to two molecules of pyruvate (steps
). Energy is then released as the two molecules of 1,3-bisphosphoglycerate are converted to two molecules of pyruvate (steps  through
through  ). Much of this energy is conserved by the coupled phosphorylation of four molecules of ADP to ATP. The net yield is two molecules of ATP per molecule of glucose used, because two molecules of ATP were invested in the preparatory phase. Energy is also conserved in the payoff phase in the formation of two molecules of the electron carrier NADH per molecule of glucose.
). Much of this energy is conserved by the coupled phosphorylation of four molecules of ADP to ATP. The net yield is two molecules of ATP per molecule of glucose used, because two molecules of ATP were invested in the preparatory phase. Energy is also conserved in the payoff phase in the formation of two molecules of the electron carrier NADH per molecule of glucose.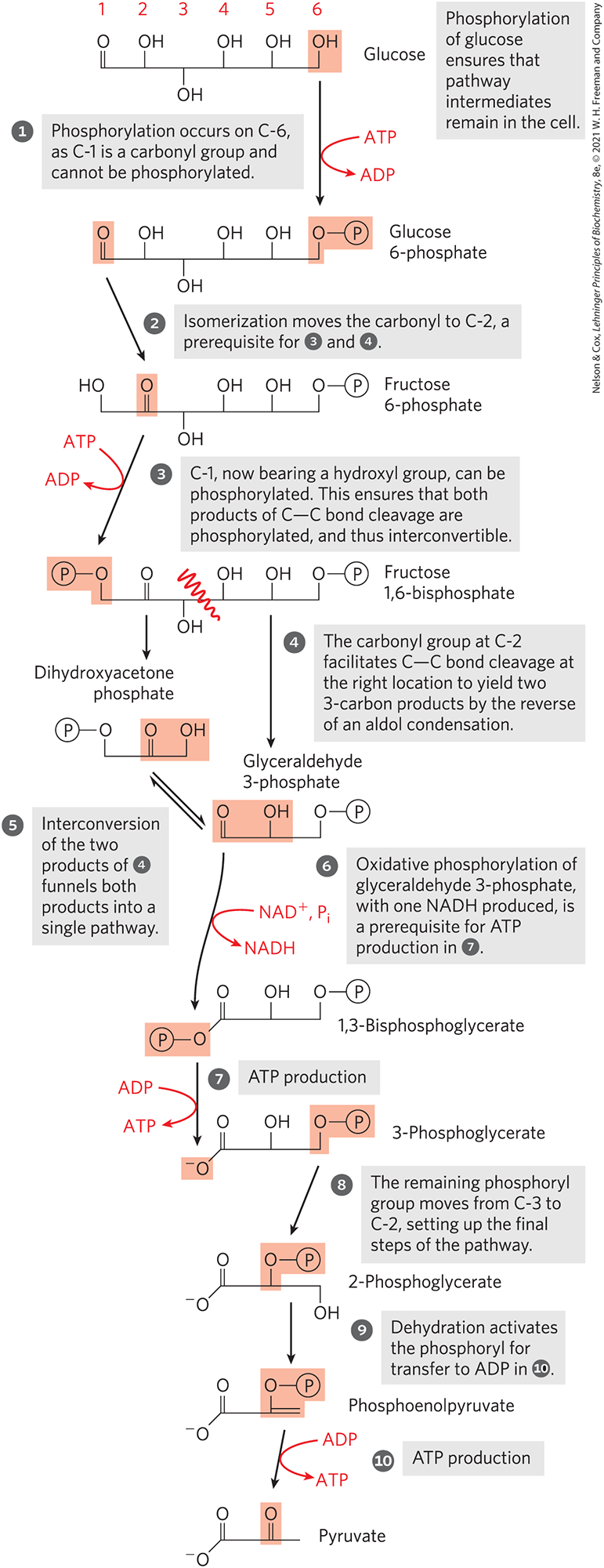

 Conversion of 3-Phosphoglycerate to 2-Phosphoglycerate
Conversion of 3-Phosphoglycerate to 2-Phosphoglycerate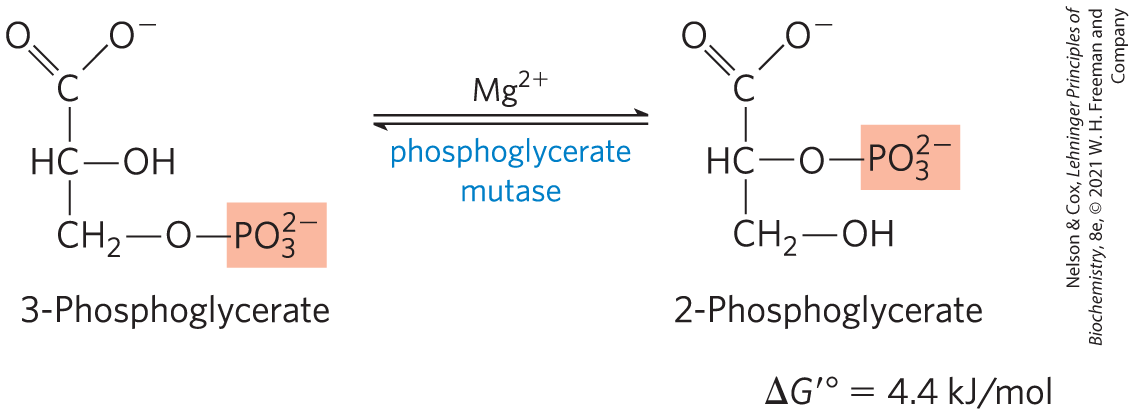
 Dehydration of 2-Phosphoglycerate to Phosphoenolpyruvate
Dehydration of 2-Phosphoglycerate to Phosphoenolpyruvate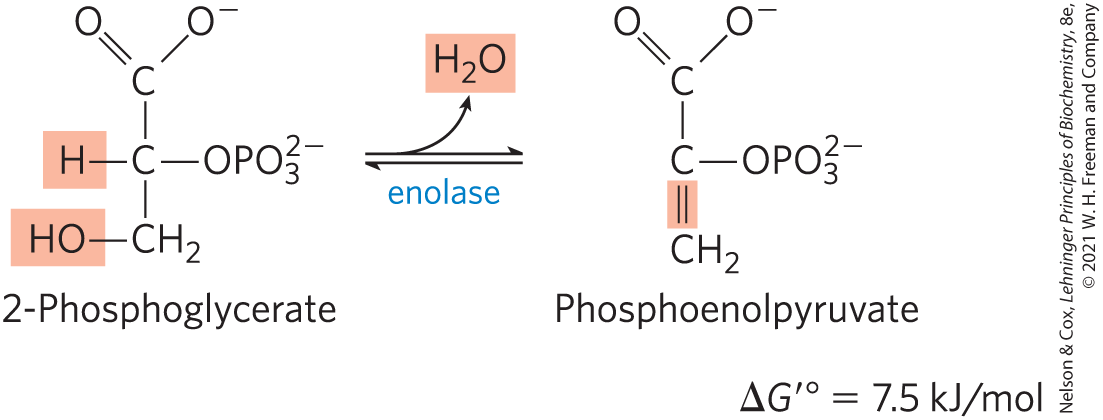
 Glycolysis is a near-universal pathway by which a glucose molecule is oxidized, in two phases, to two molecules of pyruvate, with energy conserved as ATP and NADH. Ten cytosolic enzymes act sequentially in glycolysis. The overall reaction converts glucose to two molecules of pyruvate, and energy is conserved in the synthesis of two molecules of ATP and two molecules of NADH.
Glycolysis is a near-universal pathway by which a glucose molecule is oxidized, in two phases, to two molecules of pyruvate, with energy conserved as ATP and NADH. Ten cytosolic enzymes act sequentially in glycolysis. The overall reaction converts glucose to two molecules of pyruvate, and energy is conserved in the synthesis of two molecules of ATP and two molecules of NADH.