14.6 Pentose Phosphate Pathway of Glucose Oxidation
In most animal tissues, the major catabolic fate of glucose 6-phosphate is glycolytic breakdown to pyruvate, much of which is then oxidized via the citric acid cycle, ultimately leading to the formation of ATP. Glucose 6-phosphate does have other catabolic fates, however, which lead to specialized products needed by the cell. Of particular importance in some tissues is the oxidation of glucose 6-phosphate to pentose phosphates by the pentose phosphate pathway (also called the phosphogluconate pathway or the hexose monophosphate pathway; Fig. 14-29). In this oxidative pathway, is the electron acceptor, yielding NADPH. Rapidly dividing cells, such as those of bone marrow, skin, and intestinal mucosa, and those of tumors, use the pentose ribose 5-phosphate to make RNA, DNA, and such coenzymes as ATP, NADH, , and coenzyme A.
FIGURE 14-29 General scheme of the pentose phosphate pathway. NADPH formed in the oxidative phase is used to reduce glutathione, GSSG (see Box 14-4), and to support reductive biosynthesis. The other product of the oxidative phase is ribose 5-phosphate, which serves as a precursor for nucleotides, coenzymes, and nucleic acids. In cells that are not using ribose 5-phosphate for biosynthesis, the nonoxidative phase recycles six molecules of the pentose into five molecules of the hexose glucose 6-phosphate, allowing continued production of NADPH and converting glucose 6-phosphate (in six cycles) to .
In other tissues, the essential product of the pentose phosphate pathway is not the pentoses but the electron donor NADPH, needed for reductive biosynthesis or to counter the damaging effects of oxygen radicals. Tissues that carry out extensive fatty acid synthesis (liver, adipose, lactating mammary gland) or very active synthesis of cholesterol and steroid hormones (liver, adrenal glands, gonads) require the NADPH provided by this pathway. Erythrocytes and the cells of the lens and cornea are directly exposed to oxygen and thus to the damaging free radicals generated by oxygen. By maintaining a reducing environment (a high ratio of NADPH to and a high ratio of reduced glutathione to oxidized glutathione), such cells can prevent or undo oxidative damage to proteins, lipids, and other sensitive molecules. In erythrocytes, the NADPH produced by the pentose phosphate pathway is so important in preventing oxidative damage that a genetic defect in glucose 6-phosphate dehydrogenase, the first enzyme of the pathway, can have serious medical consequences (Box 14-4).
The Oxidative Phase Produces NADPH and Pentose Phosphates
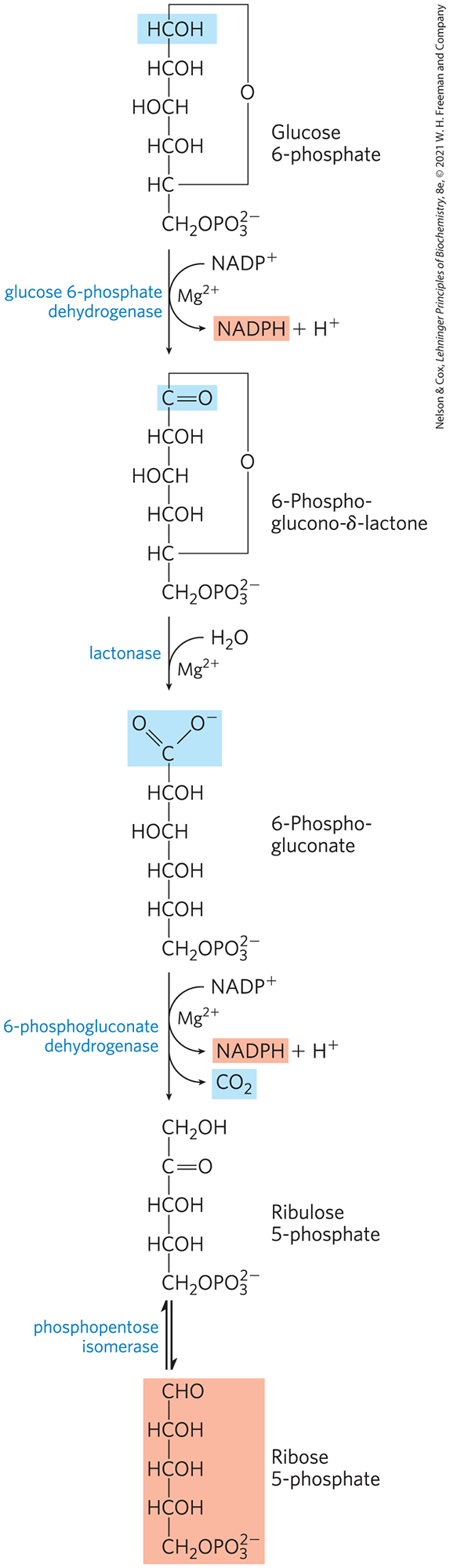
The first reaction of the pentose phosphate pathway (Fig. 14-30) is the oxidation of glucose 6-phosphate by glucose 6-phosphate dehydrogenase (G6PD) to form 6-phosphoglucono-δ-lactone, an intramolecular ester. is the electron acceptor, and the overall equilibrium lies far in the direction of NADPH formation. The lactone is hydrolyzed to the free acid 6-phosphogluconate by a specific lactonase, then 6-phosphogluconate undergoes oxidation and decarboxylation by 6-phosphogluconate dehydrogenase to form the ketopentose ribulose 5-phosphate; the reaction also generates a second molecule of NADPH. Phosphopentose isomerase converts ribulose 5-phosphate to its aldose isomer, ribose 5-phosphate. In some tissues, the pentose phosphate pathway ends at this point, and its overall equation is
FIGURE 14-30 Oxidative reactions of the pentose phosphate pathway. The end products are ribose 5-phosphate, , and NADPH.
The net result is the production of NADPH, a reductant for biosynthetic reactions, and ribose 5-phosphate, a precursor for nucleotide synthesis.
The Nonoxidative Phase Recycles Pentose Phosphates to Glucose 6-Phosphate
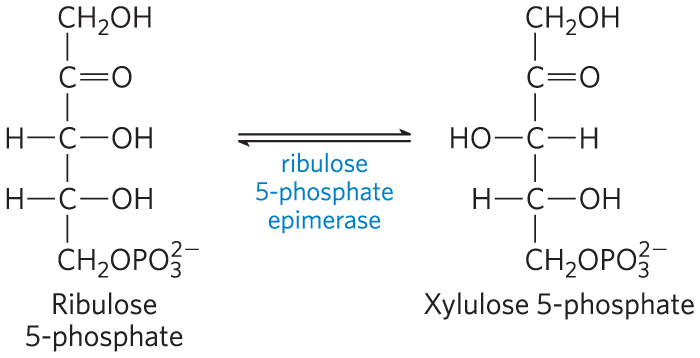
In tissues that require primarily NADPH, the pentose phosphates produced in the oxidative phase of the pathway are recycled into glucose 6-phosphate. In this nonoxidative phase, ribulose 5-phosphate is first epimerized to xylulose 5-phosphate:
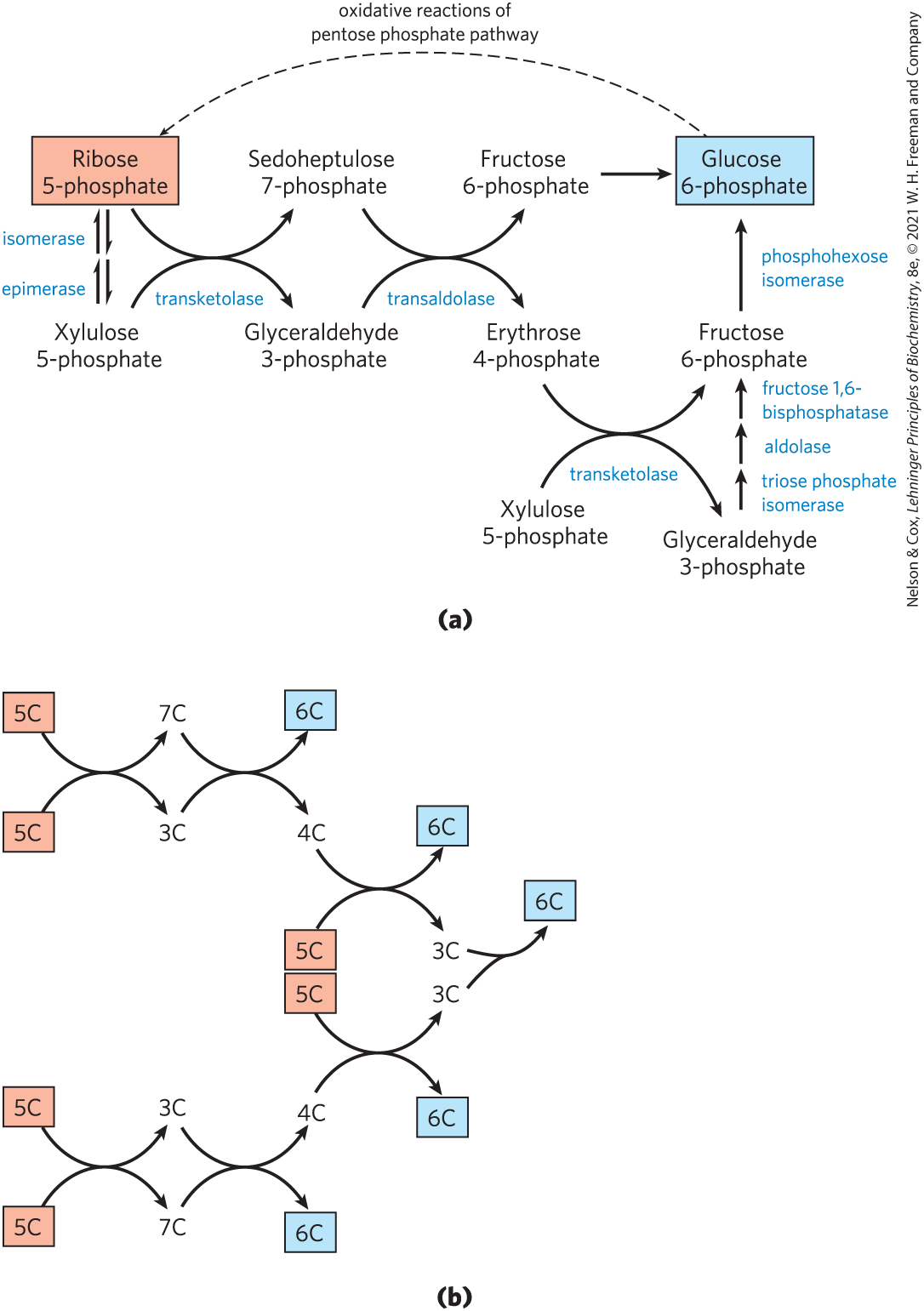
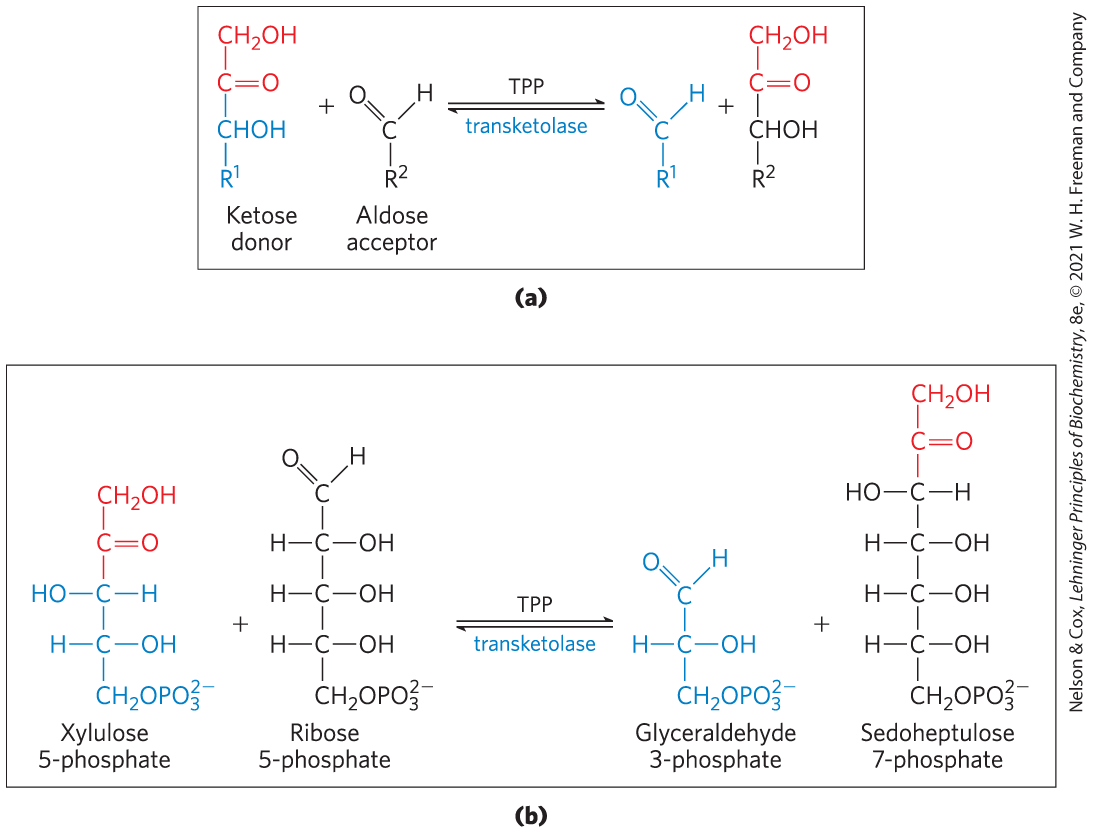
Then, in a series of rearrangements of the carbon skeletons (Fig. 14-31), six five-carbon sugar phosphates are converted to five six-carbon sugar phosphates, completing the cycle and allowing continued oxidation of glucose 6-phosphate with production of NADPH. Continued recycling leads ultimately to the conversion of glucose 6-phosphate to six . Two enzymes unique to the pentose phosphate pathway act in these interconversions of sugars: transketolase and transaldolase. Transketolase catalyzes the transfer of a two-carbon fragment from a ketose donor to an aldose acceptor (Fig. 14-32a). In its first appearance in the pentose phosphate pathway, transketolase transfers C-1 and C-2 of xylulose 5-phosphate to ribose 5-phosphate, forming the seven-carbon product sedoheptulose 7-phosphate (Fig. 14-32b). The remaining three-carbon fragment from xylulose is glyceraldehyde 3-phosphate.
FIGURE 14-31 Nonoxidative reactions of the pentose phosphate pathway. (a) These reactions convert pentose phosphates to hexose phosphates, allowing the oxidative reactions to continue. Transketolase and transaldolase are specific to this pathway; the other enzymes also serve in the glycolytic or gluconeogenic pathways. (b) A schematic diagram showing the pathway from six pentoses (5C) to five hexoses (6C). Note that this involves two sets of the interconversions shown in (a). Every reaction shown here is reversible; unidirectional arrows are used only to make clear the direction of the reactions during continuous oxidation of glucose 6-phosphate. In the light-independent reactions of photosynthesis, the direction of these reactions is reversed.
FIGURE 14-32 The first reaction catalyzed by transketolase. (a) The general reaction catalyzed by transketolase is the transfer of a two-carbon group, carried temporarily on enzyme-bound TPP, from a ketose donor to an aldose acceptor. (b) Conversion of two pentose phosphates to a triose phosphate and a seven-carbon sugar phosphate, sedoheptulose 7-phosphate.
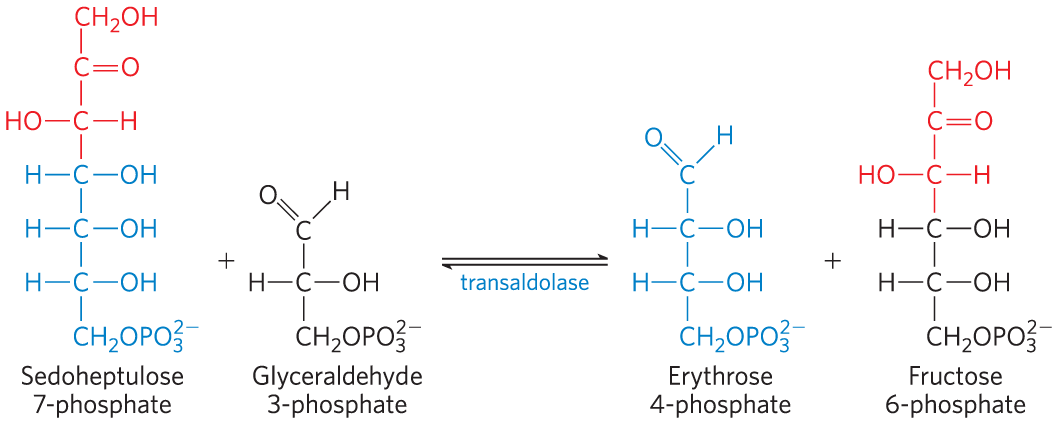
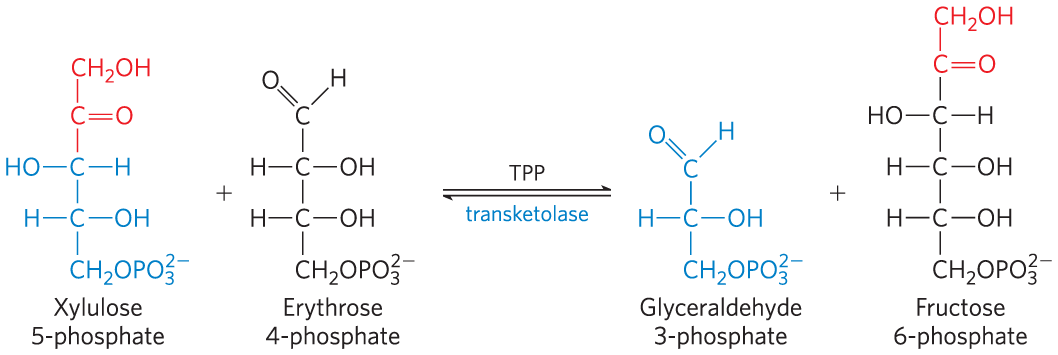
Next, transaldolase catalyzes a reaction similar to the aldolase reaction of glycolysis: a three-carbon fragment is removed from sedoheptulose 7-phosphate and condensed with glyceraldehyde 3-phosphate, forming fructose 6-phosphate and the tetrose erythrose 4-phosphate (Fig. 14-33). Now transketolase acts again, forming fructose 6-phosphate and glyceraldehyde 3-phosphate from erythrose 4-phosphate and xylulose 5-phosphate (Fig. 14-34). Two molecules of glyceraldehyde 3-phosphate formed by two iterations of these reactions can be converted to a molecule of fructose 1,6-bisphosphate as in gluconeogenesis (Fig. 14-16), and finally FBPase-1 and phosphohexose isomerase convert fructose 1,6-bisphosphate to glucose 6-phosphate. Overall, six pentose phosphates have been converted to five hexose phosphates (Fig. 14-32b) — the cycle is now complete.
FIGURE 14-33 The reaction catalyzed by transaldolase.
FIGURE 14-34 The second reaction catalyzed by transketolase.
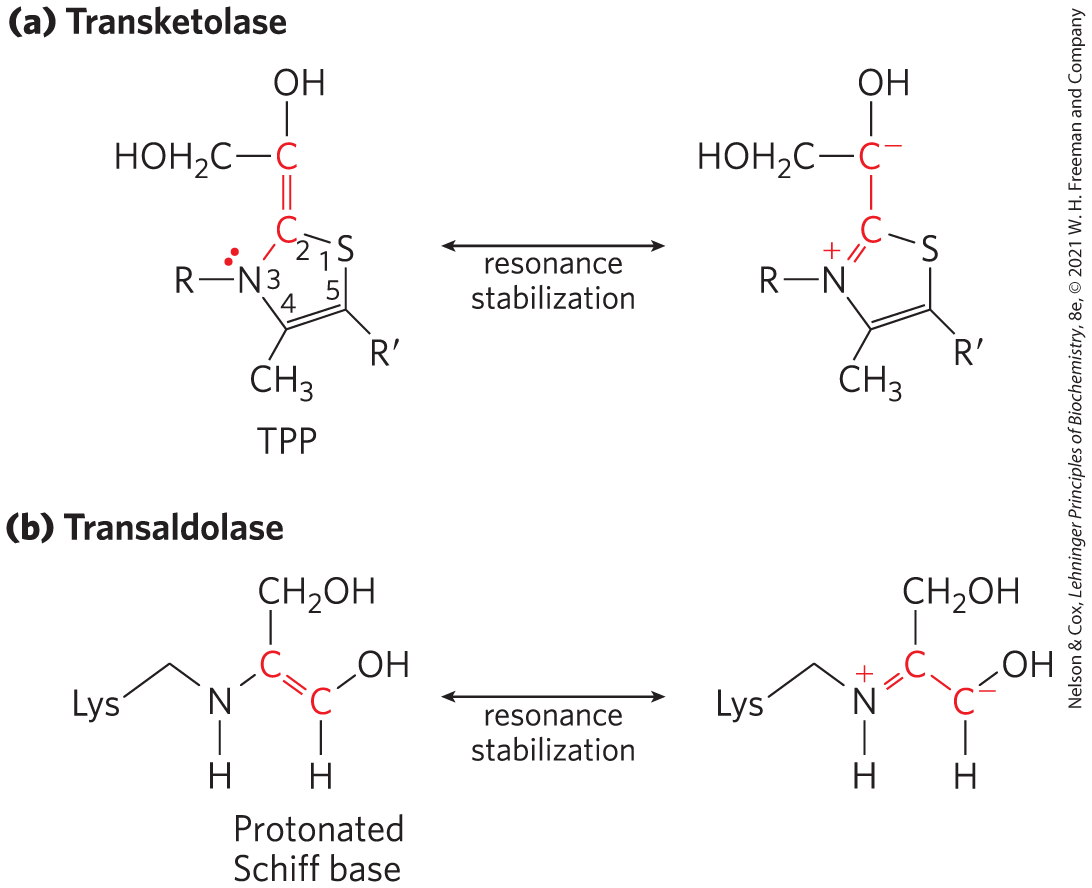
Transketolase requires the cofactor thiamine pyrophosphate (TPP), which stabilizes a two-carbon carbanion in this reaction (Fig. 14-35a), just as it does in the pyruvate decarboxylase reaction (Fig. 14-13). Transaldolase uses a Lys side chain to form a Schiff base with the carbonyl group of its substrate, a ketose, thereby stabilizing a carbanion (Fig. 14-35b) that is central to the reaction mechanism.
FIGURE 14-35 Carbanion intermediates stabilized by covalent interactions with transketolase and transaldolase. (a) The ring of TPP stabilizes the carbanion in the dihydroxyethyl group carried by transketolase. (b) In the transaldolase reaction, the protonated Schiff base formed between the ε-amino group of a Lys side chain and the substrate stabilizes the C-3 carbanion formed after aldol cleavage.
The first and third steps of the oxidative pentose phosphate pathway shown in Figure 14-30 are oxidations with large, negative standard free-energy changes and are essentially irreversible in the cell. The reactions of the nonoxidative part of the pentose phosphate pathway (Fig. 14-31) are readily reversible and thus also provide a means of converting hexose phosphates to pentose phosphates. As we shall see in Chapter 20, a process that converts hexose phosphates to pentose phosphates is central to the photosynthetic assimilation of by plants. That pathway, the reductive pentose phosphate pathway, is essentially the reversal of the reactions shown in Figure 14-31 and employs many of the same enzymes.
All the enzymes of the pentose phosphate pathway are located in the cytosol, like those of glycolysis and most of those of gluconeogenesis. In fact, these three pathways are connected through several shared intermediates and enzymes. The glyceraldehyde 3-phosphate formed by the action of transketolase is readily converted to dihydroxyacetone phosphate by the glycolytic enzyme triose phosphate isomerase, and these two trioses can be joined by the aldolase as in gluconeogenesis, forming fructose 1,6-bisphosphate. Alternatively, the triose phosphates can be oxidized to pyruvate by the glycolytic reactions. The fate of the trioses is determined by the cell’s relative needs for pentose phosphates, NADPH, and ATP.
Glucose 6-Phosphate Is Partitioned between Glycolysis and the Pentose Phosphate Pathway
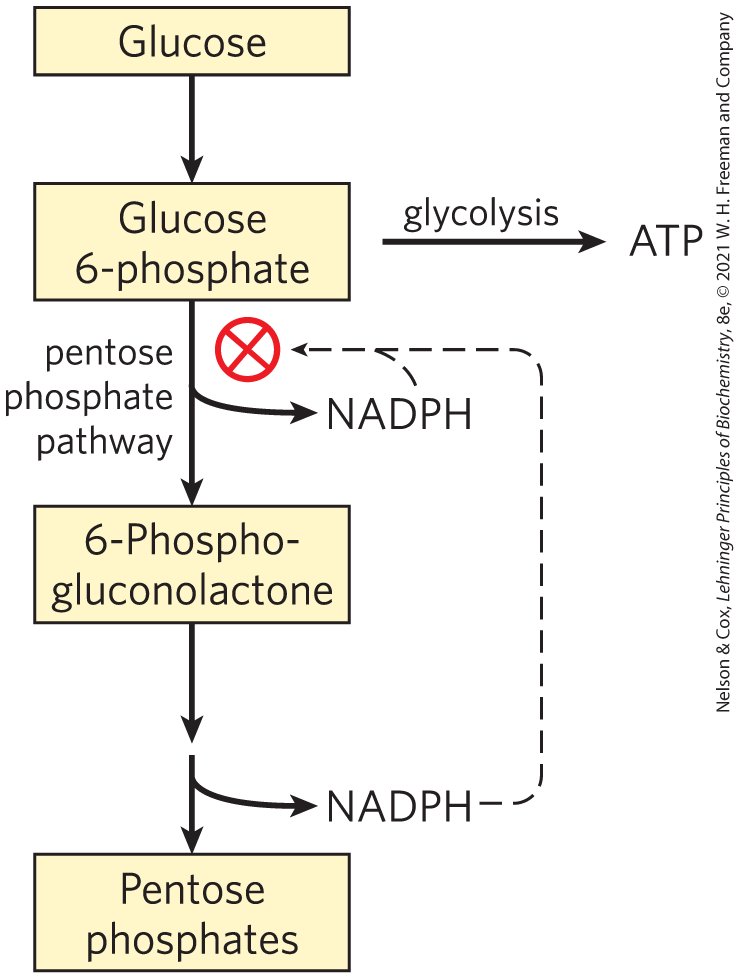
Whether glucose 6-phosphate enters glycolysis or the pentose phosphate pathway depends on the current needs of the cell and on the concentration of in the cytosol. Without this electron acceptor, the first reaction of the pentose phosphate pathway (catalyzed by glucose 6-phosphate dehydrogenase) cannot proceed. When a cell is rapidly converting NADPH to in biosynthetic reductions, rises, allosterically stimulating glucose 6-phosphate dehydrogenase and thereby increasing the flux of glucose 6-phosphate through the pentose phosphate pathway (Fig. 14-36). When the demand for NADPH slows, the level of drops, the pentose phosphate pathway slows, and glucose 6-phosphate is instead used to fuel glycolysis.
FIGURE 14-36 Role of NADPH in regulating the partitioning of glucose 6-phosphate between glycolysis and the pentose phosphate pathway. When NADPH is forming faster than it is being used for biosynthesis and glutathione reduction, [NADPH] rises and inhibits the first enzyme in the pentose phosphate pathway. As a result, more glucose 6-phosphate is available for glycolysis.
Thiamine Deficiency Causes Beriberi and Wernicke-Korsakoff Syndrome
Thiamine, precursor to the cofactor thiamine pyrophosphate (TPP), is one of the B vitamins, essential in humans. Lack of vitamin in the diet leads to a range of medical problems. The condition known as beriberi is characterized by an accumulation of body fluids (swelling), pain, paralysis, and ultimately, without treatment, death. Wernicke-Korsakoff syndrome, also caused by a severe deficiency of thiamine, typically includes problems with voluntary movements, reflected in abnormal eye movements and gait, and neurological defects. The syndrome is more common among heavy drinkers than in the general population because chronic, heavy alcohol consumption interferes with the intestinal absorption of thiamine. The syndrome can be exacerbated by a mutation in the gene for transketolase that results in an enzyme with a lowered affinity for TPP — an affinity one-tenth that of the normal enzyme. This defect makes individuals much more sensitive to a thiamine deficiency: even a moderate thiamine deficiency (tolerable in individuals with an unmutated transketolase) can result in a transketolase that is not saturated with TPP at its normal concentration. The result is a slowing down of the whole pentose phosphate pathway. In people with Wernicke-Korsakoff syndrome, this mutation results in a worsening of symptoms, which can include severe memory loss, mental confusion, and partial paralysis.
SUMMARY 14.6 Pentose Phosphate Pathway of Glucose Oxidation
The oxidative pentose phosphate pathway produces NADPH and pentose phosphates. Tissues that carry out extensive fatty acid synthesis (liver, adipose, lactating mammary gland) or very active synthesis of cholesterol and steroid hormones (liver, adrenal glands, gonads) require the NADPH provided by this pathway.
Ribose 5-phosphate is a precursor for nucleotide and nucleic acid synthesis.
The first, oxidative phase of the pentose phosphate pathway consists of two oxidations that convert glucose 6-phosphate to ribulose 5-phosphate and reduce to NADPH.
The second, nonoxidative phase of the pentose phosphate pathway comprises steps that convert pentose phosphates to glucose 6-phosphate, which begins the oxidative cycle again.
Entry of glucose 6-phosphate either into glycolysis or into the pentose phosphate pathway is largely determined by the relative concentrations of and NADPH.
 In most animal tissues, the major catabolic fate of glucose 6-phosphate is glycolytic breakdown to pyruvate, much of which is then oxidized via the citric acid cycle, ultimately leading to the formation of ATP. Glucose 6-phosphate does have other catabolic fates, however, which lead to specialized products needed by the cell. Of particular importance in some tissues is the oxidation of glucose 6-phosphate to pentose phosphates by the
In most animal tissues, the major catabolic fate of glucose 6-phosphate is glycolytic breakdown to pyruvate, much of which is then oxidized via the citric acid cycle, ultimately leading to the formation of ATP. Glucose 6-phosphate does have other catabolic fates, however, which lead to specialized products needed by the cell. Of particular importance in some tissues is the oxidation of glucose 6-phosphate to pentose phosphates by the 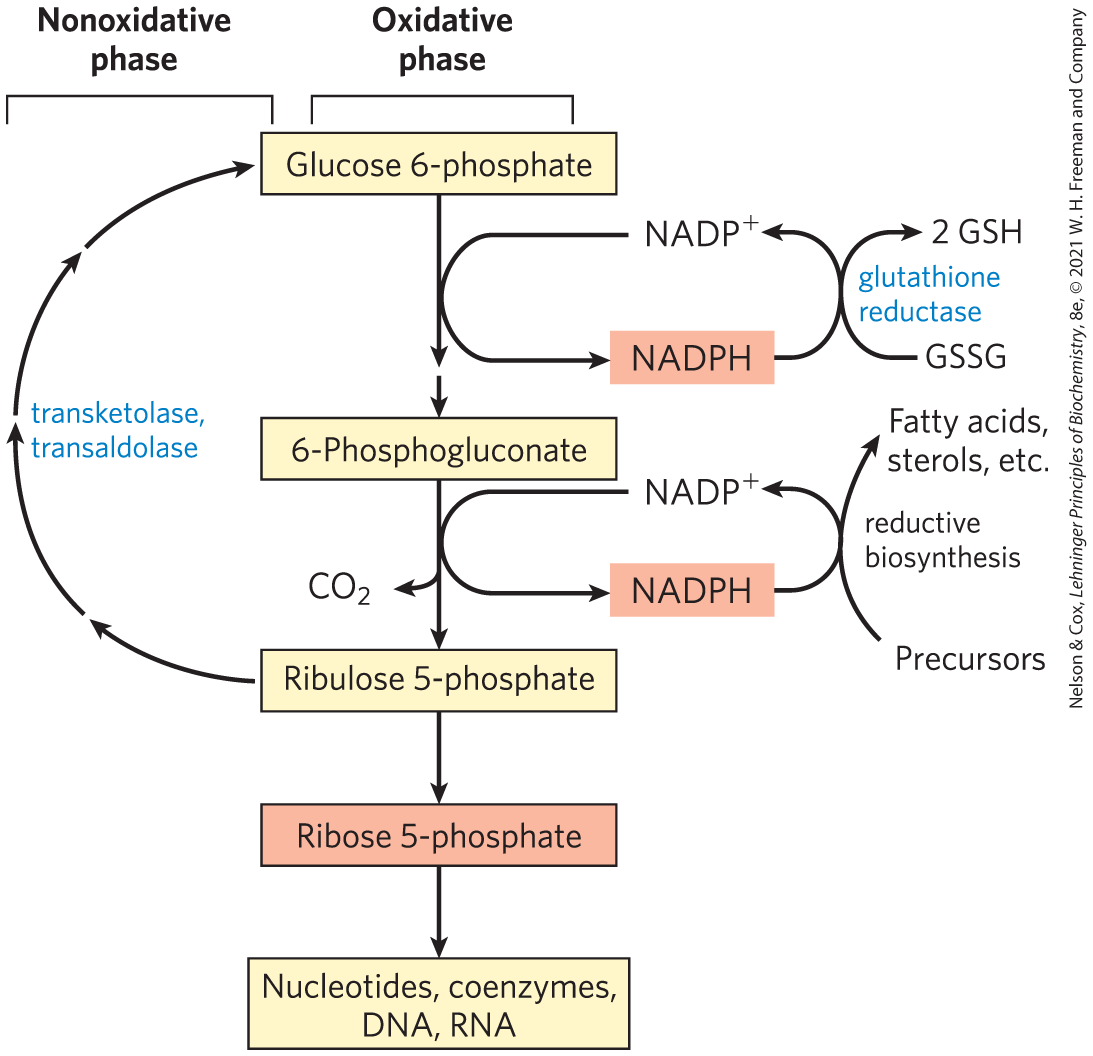
 Thiamine, precursor to the cofactor thiamine pyrophosphate (TPP), is one of the B vitamins, essential in humans. Lack of vitamin in the diet leads to a range of medical problems. The condition known as beriberi is characterized by an accumulation of body fluids (swelling), pain, paralysis, and ultimately, without treatment, death. Wernicke-Korsakoff syndrome, also caused by a severe deficiency of thiamine, typically includes problems with voluntary movements, reflected in abnormal eye movements and gait, and neurological defects. The syndrome is more common among heavy drinkers than in the general population because chronic, heavy alcohol consumption interferes with the intestinal absorption of thiamine. The syndrome can be exacerbated by a mutation in the gene for transketolase that results in an enzyme with a lowered affinity for TPP — an affinity one-tenth that of the normal enzyme. This defect makes individuals much more sensitive to a thiamine deficiency: even a moderate thiamine deficiency (tolerable in individuals with an unmutated transketolase) can result in a transketolase that is not saturated with TPP at its normal concentration. The result is a slowing down of the whole pentose phosphate pathway. In people with Wernicke-Korsakoff syndrome, this mutation results in a worsening of symptoms, which can include severe memory loss, mental confusion, and partial paralysis.
Thiamine, precursor to the cofactor thiamine pyrophosphate (TPP), is one of the B vitamins, essential in humans. Lack of vitamin in the diet leads to a range of medical problems. The condition known as beriberi is characterized by an accumulation of body fluids (swelling), pain, paralysis, and ultimately, without treatment, death. Wernicke-Korsakoff syndrome, also caused by a severe deficiency of thiamine, typically includes problems with voluntary movements, reflected in abnormal eye movements and gait, and neurological defects. The syndrome is more common among heavy drinkers than in the general population because chronic, heavy alcohol consumption interferes with the intestinal absorption of thiamine. The syndrome can be exacerbated by a mutation in the gene for transketolase that results in an enzyme with a lowered affinity for TPP — an affinity one-tenth that of the normal enzyme. This defect makes individuals much more sensitive to a thiamine deficiency: even a moderate thiamine deficiency (tolerable in individuals with an unmutated transketolase) can result in a transketolase that is not saturated with TPP at its normal concentration. The result is a slowing down of the whole pentose phosphate pathway. In people with Wernicke-Korsakoff syndrome, this mutation results in a worsening of symptoms, which can include severe memory loss, mental confusion, and partial paralysis. 
 The oxidative pentose phosphate pathway produces NADPH and pentose phosphates. Tissues that carry out extensive fatty acid synthesis (liver, adipose, lactating mammary gland) or very active synthesis of cholesterol and steroid hormones (liver, adrenal glands, gonads) require the NADPH provided by this pathway.
The oxidative pentose phosphate pathway produces NADPH and pentose phosphates. Tissues that carry out extensive fatty acid synthesis (liver, adipose, lactating mammary gland) or very active synthesis of cholesterol and steroid hormones (liver, adrenal glands, gonads) require the NADPH provided by this pathway.