Photosynthetic organisms use the ATP and NADPH produced in the light-dependent reactions of photosynthesis to synthesize all of the thousands of components that make up the organism. Plants (and other autotrophs) can reduce atmospheric to trioses, then use the trioses as precursors for the synthesis of sucrose and starch, lipids and proteins, and the many other organic components of plant cells (Fig. 20-25). Lacking these synthetic capacities, humans and other animals are ultimately dependent on photosynthetic organisms to provide the reduced fuels and organic precursors essential to life.
FIGURE 20-25 Products of photosynthesis.
Green plants contain in their chloroplasts the enzymatic machinery that catalyzes the conversion of to simple (reduced) organic compounds, a process called assimilation. This process has also been called fixation, but we reserve this term for the specific reaction in which is incorporated (fixed) into a three-carbon organic compound, the triose phosphate 3-phosphoglycerate. This simple product of photosynthesis is the precursor of more-complex biomolecules, including sugars, polysaccharides, and the metabolites derived from them, all of which are synthesized by metabolic pathways similar to those of animal tissues. Carbon dioxide is assimilated via a cyclic pathway, its key intermediates constantly regenerated. The pathway was elucidated in the early 1950s by Melvin Calvin, Andrew Benson, and James A. Bassham and is often called the Calvin cycle or, more descriptively, the reductive pentose phosphate pathway. It is essentially the reversal of a central pathway of glucose oxidation, the pentose phosphate pathway, which we described in Section 14.6.
Carbohydrate metabolism is more complex in plant cells than in animal cells or in nonphotosynthetic microorganisms. In addition to the universal pathways of glycolysis, gluconeogenesis, and the pentose phosphate pathway, plants have the unique reaction sequences for reduction of to triose phosphates and the associated reductive pentose phosphate pathway — all of which must be coordinately regulated to ensure proper allocation of carbon to energy production and synthesis of starch and sucrose. Key enzymes are regulated, as we shall see, by (1) reduction of disulfide bonds by electrons flowing from photosystem I and (2) changes in pH and concentration that result from illumination. When we look at other aspects of plant carbohydrate metabolism, we also find enzymes that are modulated by (3) conventional allosteric regulation by one or more metabolic intermediates and (4) covalent modification (phosphorylation).
Carbon Dioxide Assimilation Occurs in Three Stages
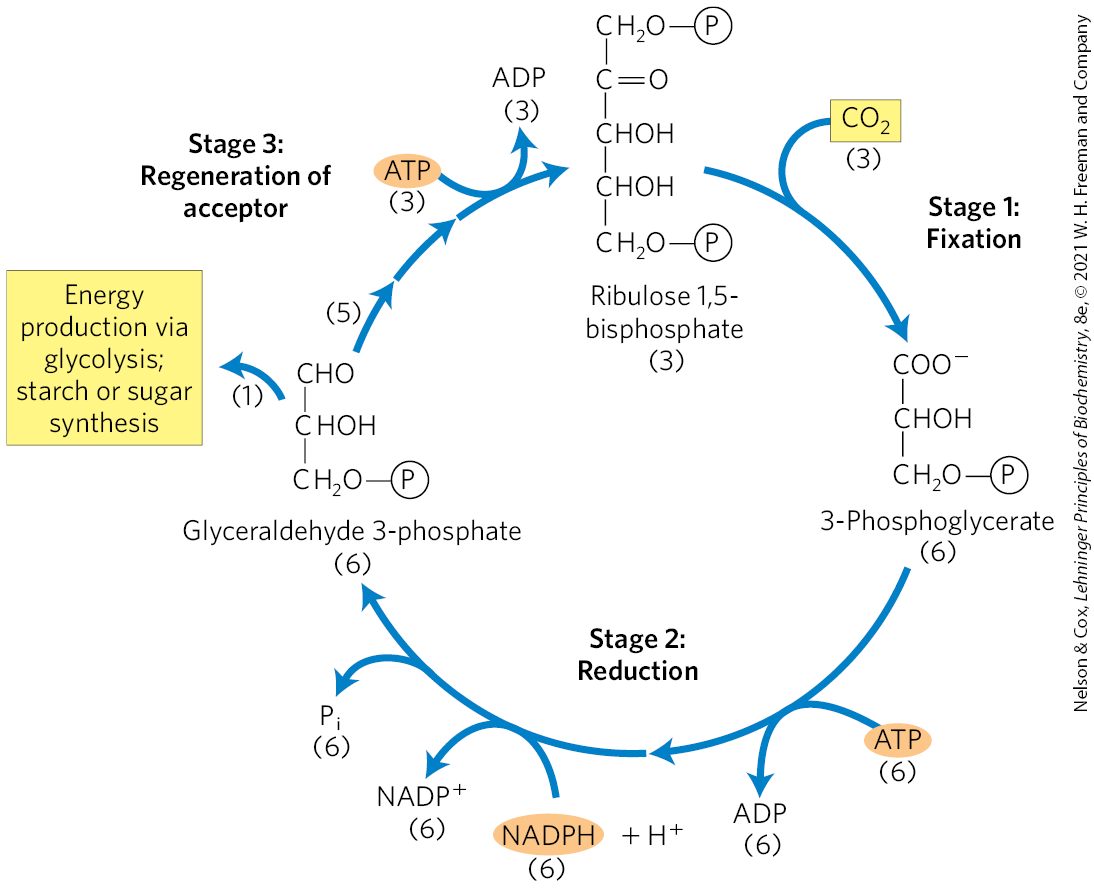
The first stage in the assimilation of into biomolecules (Fig. 20-26) is the -fixation reaction: condensation of with a five-carbon acceptor, ribulose 1,5-bisphosphate, to form two molecules of 3-phosphoglycerate. In the second stage, the 3-phosphoglycerate is reduced to triose phosphates. Overall, three molecules of are fixed to three molecules of ribulose 1,5-bisphosphate to form six molecules of glyceraldehyde 3-phosphate (18 carbons). In the third stage, five of the six molecules of triose phosphate (15 carbons) are used to regenerate three molecules of ribulose 1,5-bisphosphate (15 carbons), the starting material. The sixth molecule of triose phosphate, the net product of photosynthesis, can be used to make hexoses for fuel and building materials, sucrose for transport to nonphotosynthetic tissues, or starch for storage. Thus, the overall process is cyclical, with the continuous conversion of to triose and hexose phosphates.
FIGURE 20-26 The three stages of assimilation in photosynthetic organisms. Stoichiometries of three key intermediates (numbers in parentheses) reveal the fate of carbon atoms entering and leaving the photosynthetic carbon-reduction cycle (Calvin cycle). Three are fixed for the net synthesis of one molecule of glyceraldehyde 3-phosphate.
Fructose 6-phosphate is a key intermediate in stage 3 of assimilation; it stands at a branch point, leading either to regeneration of ribulose 1,5-bisphosphate or to synthesis of starch. The pathway from hexose phosphate to pentose bisphosphate involves many of the same reactions used in animal cells for the conversion of pentose phosphates to hexose phosphates during the nonoxidative phase of the pentose phosphate pathway (see Fig. 14-31). In the photosynthetic assimilation of , essentially the same set of reactions operates in the reverse direction, converting hexose phosphates to pentose phosphates. This reductive pentose phosphate cycle uses the same enzymes as the oxidative pathway, and several additional enzymes that make the reductive cycle irreversible. All 13 enzymes of the pathway are in the chloroplast stroma.
Stage 1: Fixation of into 3-Phosphoglycerate
An important clue to the nature of the -assimilation mechanisms in photosynthetic organisms came in the late 1940s. Calvin and his associates illuminated a suspension of green algae in the presence of radioactive carbon dioxide for just a few seconds, then quickly killed the cells, extracted their contents, and used chromatographic methods to search for the metabolites in which the labeled carbon first appeared. The first compound that became labeled was 3-phosphoglycerate, with the predominantly located in the carboxyl carbon atom. These experiments strongly suggested that 3-phosphoglycerate is an early intermediate in photosynthesis.
The many plants in which this three-carbon compound is the first intermediate are called plants, in contrast to the plants described below. Most plant species — 80% to 90% — are , including most trees, wheat, oats, rice, beans, peas, and spinach. The enzyme that catalyzes incorporation of into an organic form is ribulose 1,5-bisphosphate carboxylase/oxygenase, a name mercifully shortened to rubisco. As a carboxylase, rubisco catalyzes the covalent attachment of to the five-carbon sugar ribulose 1,5-bisphosphate and cleavage of the unstable six-carbon intermediate to form two molecules of 3-phosphoglycerate, one of which bears the carbon introduced as in its carboxyl group (Fig. 20-26). The enzyme’s oxygenase activity is discussed in Section 20.5.
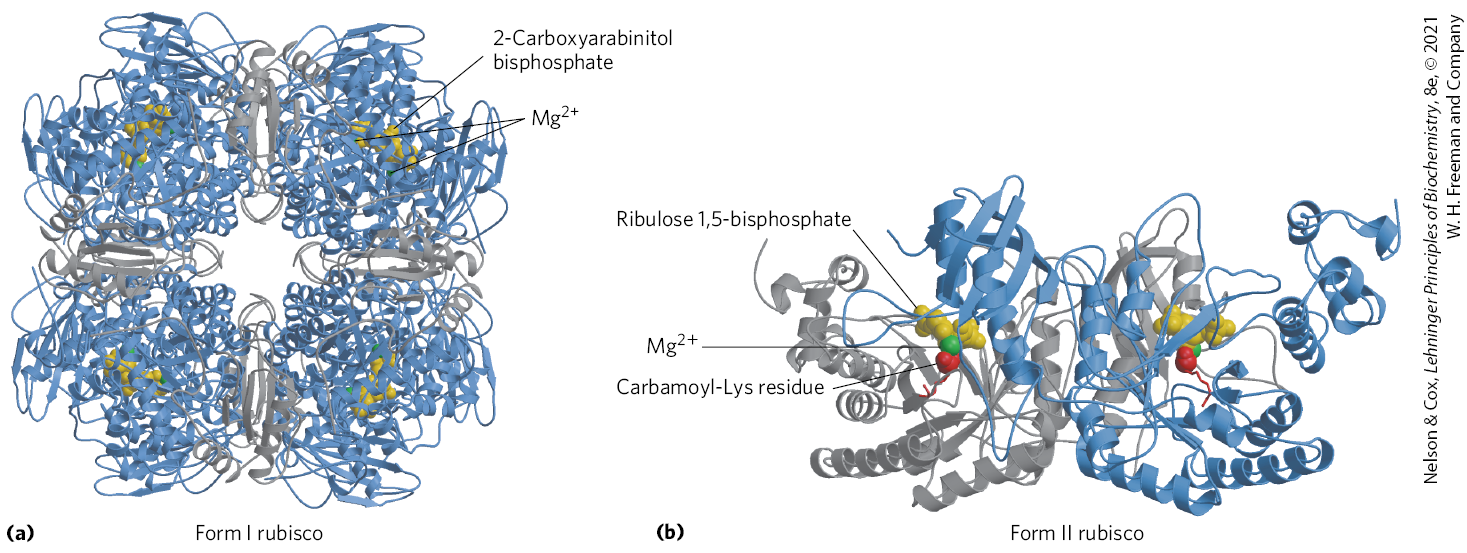
There are two distinct forms of rubisco. The rubisco of vascular plants, algae, and cyanobacteria is a crucial enzyme in the production of biomass from . It has a complex form I structure (Fig. 20-27a), with eight identical large catalytic subunits (; encoded in the chloroplast genome), and eight identical small subunits (; encoded in the nuclear genome) of uncertain function. The form II rubisco of photosynthetic bacteria is simpler, having two subunits that in many respects resemble the large subunits of the plant enzyme (Fig. 20-27b). The plant enzyme has an exceptionally low turnover number; only three molecules of are fixed per second per molecule of rubisco at 25 °C. To achieve high rates of fixation, plants therefore need large amounts of this enzyme. Rubisco is present at about 250 mg/mL in the chloroplast stroma, corresponding to an extraordinarily high concentration of active sites (~4 mm). In fact, rubisco makes up almost 50% of soluble protein in chloroplasts and is probably one of the most abundant enzymes in the biosphere.
FIGURE 20-27 Structure of ribulose 1,5-bisphosphate carboxylase/oxygenase (rubisco). (a) A ribbon model of form I rubisco from spinach. The enzyme has eight large (blue) and eight small (gray) subunits, tightly packed into a structure of . A transition-state analog, 2-carboxyarabinitol bisphosphate (yellow), is shown bound to each of the eight substrate-binding sites. in the active site is shown in green. (b) Ribbon model of form II rubisco from the bacterium Rhodospirillum rubrum. The identical subunits are in gray and blue. [Data from (a) PDB ID 8RUC, I. Andersson, J. Mol. Biol. 259:160, 1996; (b) PDB ID 9RUB, T. Lundqvist and G. Schneider, J. Biol. Chem. 266:12,604, 1991.]
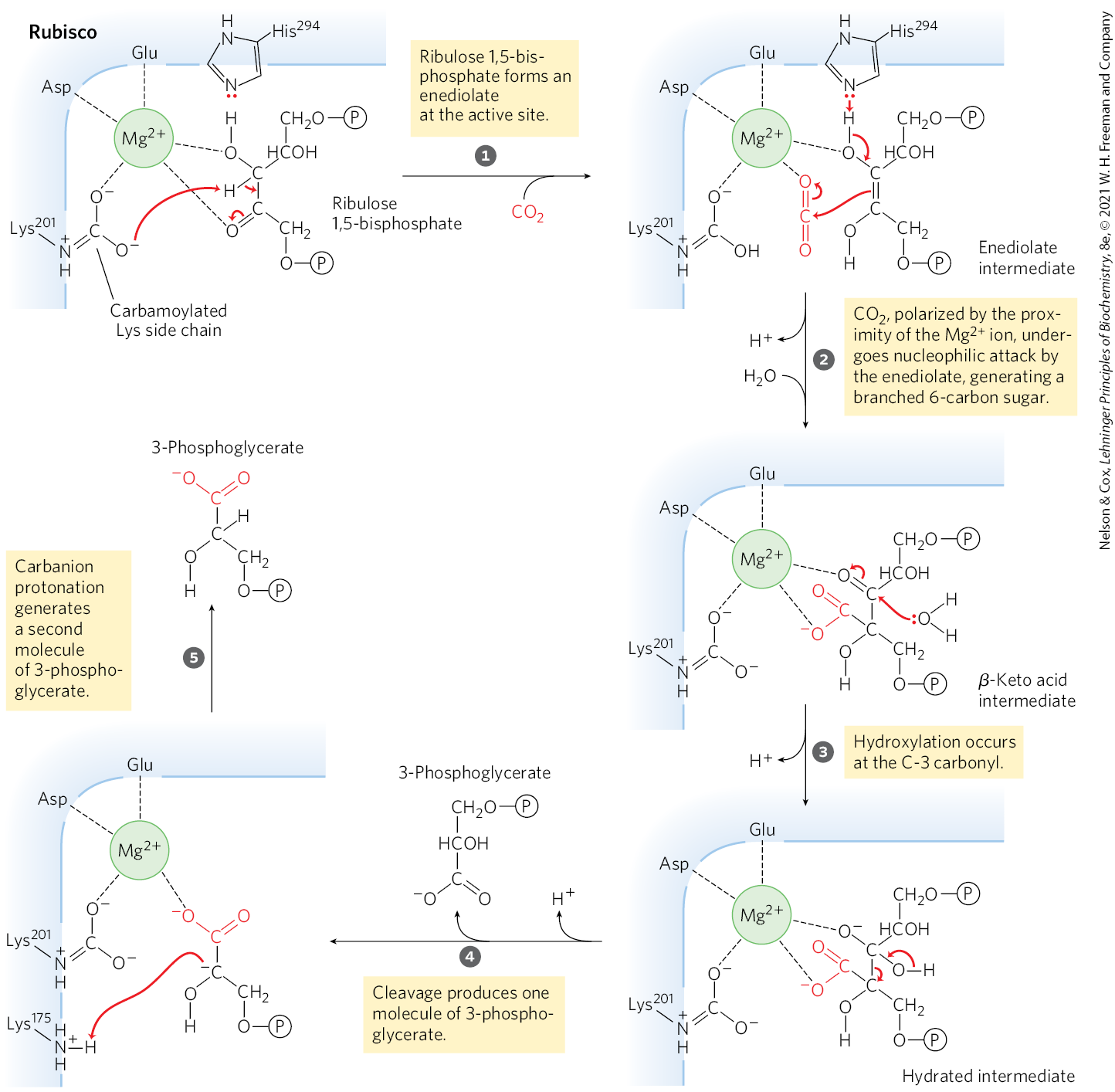
Central to the proposed mechanism for plant rubisco is a carbamoylated Lys side chain with a bound ion. The ion brings together and orients the reactants at the active site (Fig. 20-28), setting up for a nucleophilic attack by the five-carbon enediolate reaction intermediate formed on the enzyme (Fig. 20-29). The resulting six-carbon intermediate breaks down to yield two molecules of 3-phosphoglycerate.
FIGURE 20-28 Central role of in the active site of rubisco. is coordinated in a roughly octahedral complex with six oxygen atoms: one oxygen in the carbamate on ; two in the carboxyl groups of and ; two at C-2 and C-3 of the substrate, ribulose 1,5-bisphosphate; and one in the other substrate, . A water molecule occupies the -binding site in the crystal structure. In this figure, a molecule is modeled in its place. (Residue numbers refer to the spinach enzyme.) [Data from PDB ID 1RXO, T. C. Taylor and I. Andersson, J. Mol. Biol. 265:432, 1997.]
MECHANISM FIGURE 20-29 First stage of assimilation: rubisco’s carboxylase activity. The -fixation reaction is catalyzed by ribulose 1,5-bisphosphate carboxylase/oxygenase. The overall reaction accomplishes the combination of one and one ribulose 1,5-bisphosphate to form two molecules of 3-phosphoglycerate, one of which contains the carbon atom from (red). Additional proton transfers (not shown), involving , and , occur in several of these steps.
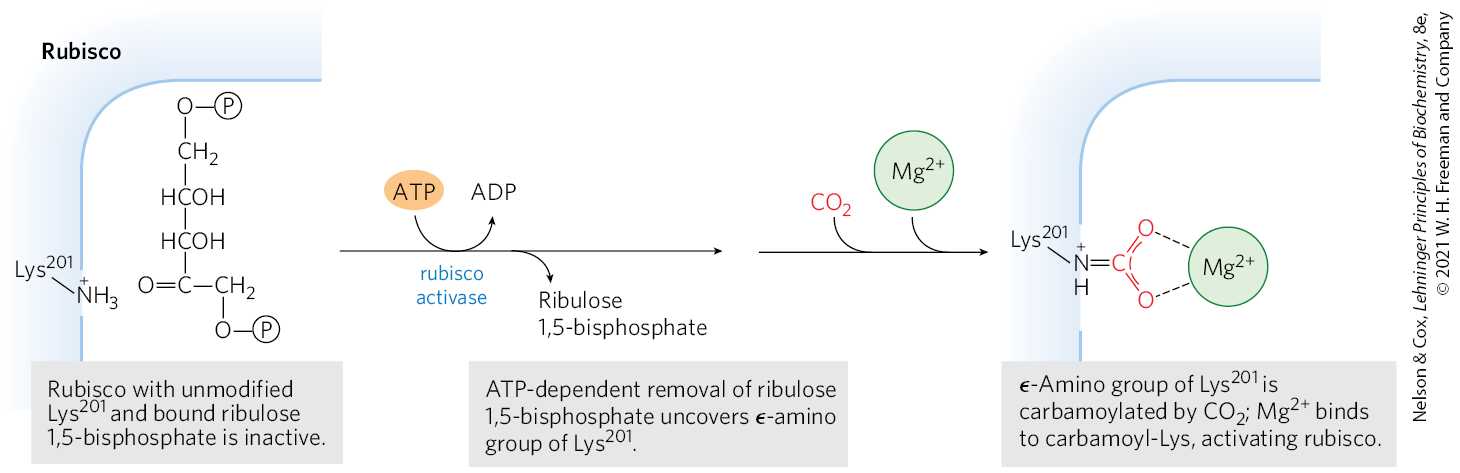
As the catalyst for the first step of photosynthetic assimilation, rubisco is a prime target for regulation. The enzyme is inactive until carbamoylated on the ε-amino group of (Fig. 20-30). Ribulose 1,5-bisphosphate inhibits carbamoylation by binding tightly to the active site and locking the enzyme in the “closed” conformation, in which is inaccessible. Rubisco activase overcomes the inhibition by promoting ATP-dependent release of the ribulose 1,5-bisphosphate, exposing the Lys amino group to nonenzymatic carbamoylation by ; this is followed by binding, which activates the rubisco.
FIGURE 20-30 Role of rubisco activase in carbamoylation of of rubisco.
Stage 2: Conversion of 3-Phosphoglycerate to Glyceraldehyde 3-Phosphate
Stage 2 begins as stromal 3-phosphoglycerate kinase catalyzes the transfer of a phosphoryl group from ATP to 3-phosphoglycerate, yielding 1,3-bisphosphoglycerate. Next, NADPH donates electrons in a reduction catalyzed by the chloroplast-specific isozyme of glyceraldehyde 3-phosphate dehydrogenase, producing glyceraldehyde 3-phosphate and . The high concentrations of NADPH and ATP in the chloroplast stroma allow this thermodynamically unfavorable pair of reactions to proceed in the direction of glyceraldehyde 3-phosphate formation. Triose phosphate isomerase then interconverts glyceraldehyde 3-phosphate and dihydroxyacetone phosphate, producing the two substrates for aldolase, which condenses them into fructose 1,6-bisphosphate. Thus far, the process has employed the same enzymes we saw in glycolysis, but operating in the reverse direction.
Most of the triose phosphate and fructose 1,6-bisphosphate produced by photosynthesis is used to regenerate ribulose 1,5-bisphosphate, the essential starting material for photosynthesis. Any excess triose phosphate is either converted to starch in the chloroplast and stored for later use or immediately exported to the cytosol and converted to sucrose for transport to growing regions of the plant.
Stage 3: Regeneration of Ribulose 1,5-Bisphosphate from Triose Phosphates
For the continuous flow of into carbohydrate, ribulose 1,5-bisphosphate must be constantly regenerated. This is accomplished in a series of mostly reversible reactions (Fig. 20-31) that, together with stages 1 and 2, constitute the reductive pentose phosphate pathway summarized in Figure 20-26. Three exergonic reactions, shown with blue arrows in Figure 20-31, make the whole process irreversible. These are the reactions catalyzed by fructose 1,6-bisphosphatase, sedoheptulose 1,7-bisphosphatase, and ribulose 5-phosphate kinase.
FIGURE 20-31 Third stage of assimilation. This schematic diagram shows the interconversions of triose phosphates and pentose phosphates. Red dots represent the number of carbons in each compound. Compounds that appear more than once are highlighted. The starting materials are glyceraldehyde 3-phosphate and dihydroxyacetone phosphate. Reactions catalyzed by aldolase ( and ) and transketolase ( and ) produce pentose phosphates that are converted to ribulose 1,5-bisphosphate — ribose 5-phosphate by ribose 5-phosphate isomerase and xylulose 5-phosphate by ribulose 5-phosphate epimerase. Ribulose 5-phosphate is phosphorylated , regenerating ribulose 1,5-bisphosphate. The reactions with blue arrows are exergonic and make the whole process irreversible: fructose 1,6-bisphosphatase, sedoheptulose 1,7-bisphosphatase, and ribulose 5-phosphate kinase.
Synthesis of Each Triose Phosphate from Requires Six NADPH and Nine ATP
The net result of three turns of the Calvin cycle is the conversion of three molecules of and one molecule of phosphate to a molecule of triose phosphate. The stoichiometry of the overall path from to triose phosphate, with regeneration of ribulose 1,5-bisphosphate, is shown in Figure 20-32.
FIGURE 20-32 Stoichiometry of assimilation in the Calvin cycle. For every three molecules fixed, one molecule of triose phosphate (glyceraldehyde 3-phosphate) is produced and nine ATP and six NADPH are consumed.
One molecule of glyceraldehyde 3-phosphate is the net product of the -assimilation pathway. The other five triose phosphate molecules (15 carbons) are rearranged in steps to of Figure 20-31 to form three molecules of ribulose 1,5-bisphosphate (15 carbons). The last step in this conversion requires one ATP per ribulose 1,5-bisphosphate, or a total of three ATP. Thus, in summary, for every molecule of triose phosphate produced by photosynthetic assimilation, six NADPH and nine ATP are required.
NADPH and ATP are produced in the light-dependent reactions of photosynthesis in about the same ratio (2:3) as they are consumed in the Calvin cycle. Nine ATP molecules are converted to ADP and phosphate in the generation of a molecule of triose phosphate; eight of the phosphates are released as and combined with eight ADP to regenerate ATP. The ninth phosphate is incorporated into the triose phosphate itself. To convert the ninth ADP to ATP, a molecule of must be imported from the cytosol, as we shall see.
In the dark, the production of ATP and NADPH by photophosphorylation and the incorporation of into triose phosphate (once referred to as the dark reactions) cease. The “dark reactions” of photosynthesis were so named to distinguish them from the primary light-driven reactions of electron transfer to and synthesis of ATP. They do not, in fact, occur at significant rates in the dark and are thus more appropriately called the -assimilation reactions. Later in this section we describe the regulatory mechanisms that turn assimilation on in the light and turn it off in the dark.
The chloroplast stroma contains all the enzymes necessary to convert the triose phosphates produced by assimilation (glyceraldehyde 3-phosphate and dihydroxyacetone phosphate) to starch, which is temporarily stored in the chloroplast as insoluble granules. Aldolase condenses the triose phosphates to fructose 1,6-bisphosphate; fructose 1,6-bisphosphatase produces fructose 6-phosphate; phosphohexose isomerase yields glucose 6-phosphate; and phosphoglucomutase produces glucose 1-phosphate, the starting material for starch synthesis (see Section 20.6).
All the reactions of the Calvin cycle except those catalyzed by rubisco, sedoheptulose 1,7-bisphosphatase, and ribulose 5-phosphate kinase also take place in animal tissues. Lacking these three enzymes, animals cannot carry out significant conversion of to glucose.
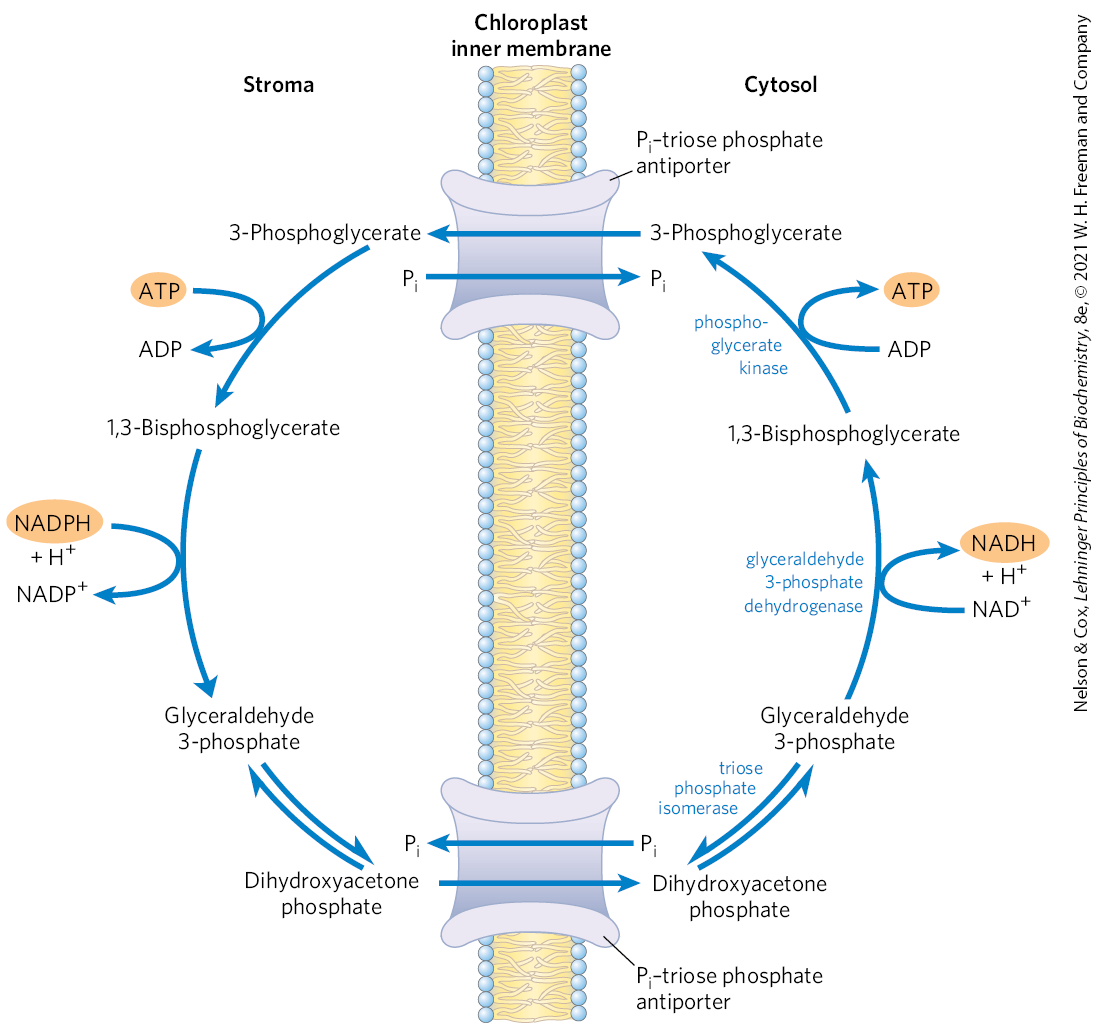
A Transport System Exports Triose Phosphates from the Chloroplast and Imports Phosphate
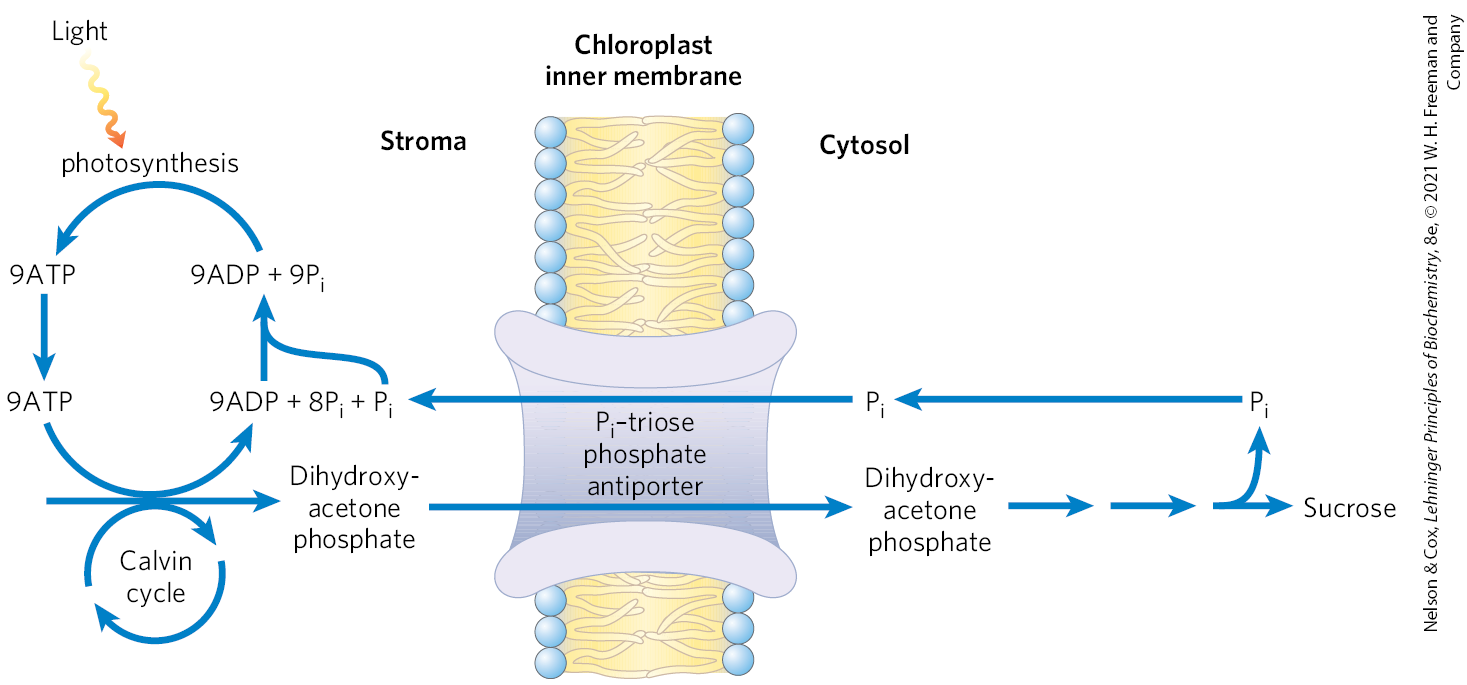
The inner chloroplast membrane is impermeable to most phosphorylated compounds, including fructose 6-phosphate, glucose 6-phosphate, and fructose 1,6-bisphosphate. It does, however, have a specific antiporter that catalyzes the one-for-one exchange of with a triose phosphate, either dihydroxyacetone phosphate or 3-phosphoglycerate (Fig. 20-33). This antiporter simultaneously moves into the chloroplast, where it is used in photophosphorylation, and moves triose phosphate into the cytosol, where it can be used to synthesize sucrose, the form in which the fixed carbon is transported to distant plant tissues.
FIGURE 20-33 The phosphate antiport system of the chloroplast inner membrane. This transporter facilitates the exchange of cytosolic for stromal dihydroxyacetone phosphate. The products of photosynthetic assimilation are thus moved into the cytosol, where they serve as a starting point for sucrose biosynthesis, and required for photophosphorylation is moved into the stroma. This same antiporter can transport 3-phosphoglycerate, and it acts indirectly in the export of ATP and reducing equivalents (see Fig. 20-34).
Sucrose synthesis in the cytosol and starch synthesis in the chloroplast are the major pathways by which the excess triose phosphate from photosynthesis is harvested. Sucrose synthesis (described later) releases four molecules from the four triose phosphates required to make sucrose. For every molecule of triose phosphate removed from the chloroplast, one is transported into the chloroplast, providing the ninth mentioned above, to be used in regenerating ATP. If this exchange were blocked, triose phosphate synthesis would quickly deplete the available in the chloroplast, slowing ATP synthesis and suppressing assimilation of into starch.
The –triose phosphate antiport system serves one additional function. ATP and reducing power are needed in the cytosol for a variety of synthetic and energy-requiring reactions. These requirements are met to an as-yet-undetermined degree by mitochondria, but a second potential source of energy is the ATP and NADPH generated in the chloroplast stroma during the light-dependent reactions. However, neither ATP nor NADPH can cross the chloroplast membrane. The –triose phosphate antiport system has the indirect effect of moving ATP equivalents and reducing equivalents from the chloroplast to the cytosol (Fig. 20-34). Dihydroxyacetone phosphate formed in the stroma is transported to the cytosol, where it is converted by glycolytic enzymes to 3-phosphoglycerate, generating ATP and NADH. 3-Phosphoglycerate reenters the chloroplast, completing the cycle.
FIGURE 20-34 Role of the phosphate antiporter in the transport of ATP and reducing equivalents. Dihydroxyacetone phosphate leaves the chloroplast and is converted to glyceraldehyde 3-phosphate in the cytosol. The cytosolic glyceraldehyde 3-phosphate dehydrogenase and phosphoglycerate kinase reactions then produce NADH, ATP, and 3-phosphoglycerate. The latter reenters the chloroplast and is reduced to dihydroxyacetone phosphate, completing a cycle that effectively moves ATP and reducing equivalents (NAD(P)H) from chloroplast to cytosol.
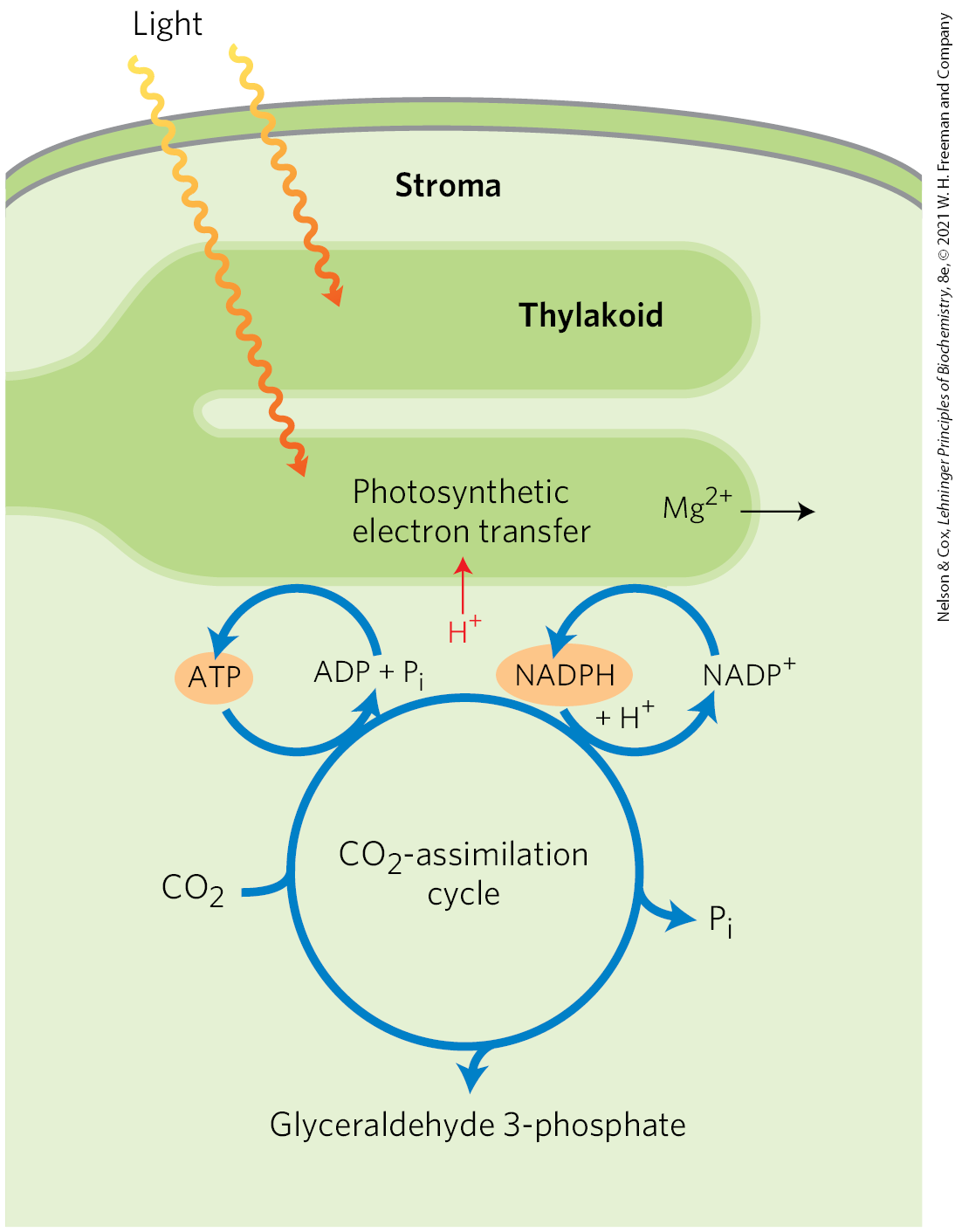
Four Enzymes of the Calvin Cycle Are Indirectly Activated by Light
The reductive assimilation of requires a lot of ATP and NADPH, and their stromal concentrations increase when chloroplasts are illuminated (Fig. 20-35). The light-induced transport of protons across the thylakoid membrane also increases the stromal pH from about 7 to about 8, and it is accompanied by a flow of from the thylakoid compartment into the stroma, raising the from 1 to 3 mm to 3 to 6 mm. Several stromal enzymes have evolved to take advantage of these light-induced conditions, which signal the availability of ATP and NADPH: the enzymes are more active in an alkaline environment and at high . For example, activation of rubisco by formation of carbamoyllysine is faster at alkaline pH, and high stromal favors formation of the enzyme’s active complex. Fructose 1,6-bisphosphatase requires and is very dependent on pH (Fig. 20-36); its activity increases more than 100-fold when pH and rise during chloroplast illumination.
FIGURE 20-35 Source of ATP and NADPH. ATP and NADPH produced by the light-dependent reactions are essential substrates for the reduction of . The photosynthetic reactions that produce ATP and NADPH are accompanied by movement of protons (red) from the stroma into the thylakoid, creating alkaline conditions in the stroma. Magnesium ions pass from the thylakoid into the stroma, increasing the stromal .
FIGURE 20-36 Activation of chloroplast fructose 1,6-bisphosphatase. Reduced fructose 1,6-bisphosphatase (FBPase-1) is activated by light and by the combination of high pH and high in the stroma, both of which are results of illumination. [Information from B. Halliwell, Chloroplast Metabolism: The Structure and Function of Chloroplasts in Green Leaf Cells, p. 97, Clarendon Press, 1984.]
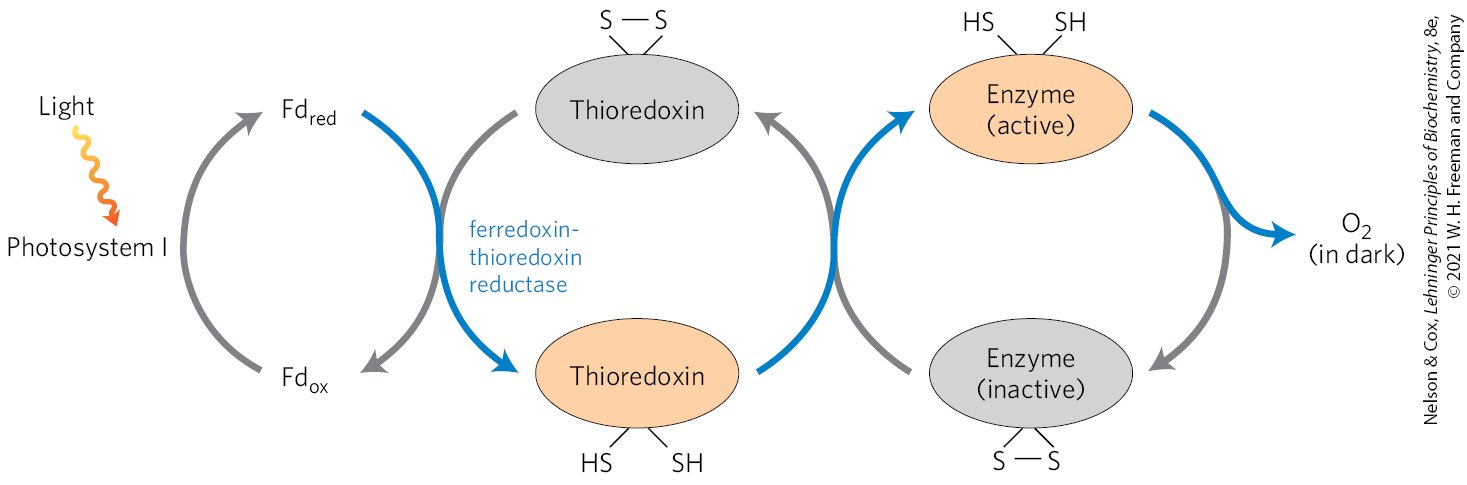
Four Calvin cycle enzymes are subject to a special type of regulation by light. Ribulose 5-phosphate kinase, fructose 1,6-bisphosphatase, sedoheptulose 1,7-bisphosphatase, and glyceraldehyde 3-phosphate dehydrogenase are activated by light-driven reduction of disulfide bonds between two Cys residues critical to their catalytic activities. When these Cys residues are disulfide-bonded (oxidized), the enzymes are inactive; this is the normal situation in the dark. With illumination, electrons flow from photosystem I to ferredoxin (Fig. 20-12), which passes electrons to a small, soluble, disulfide-containing protein called thioredoxin (Fig. 20-37), in a reaction catalyzed by ferredoxin: thioredoxin reductase. Reduced thioredoxin donates electrons for the reduction of the disulfide bonds of the light-activated enzymes, and these reductive cleavage reactions are accompanied by conformational changes that increase enzyme activities. At nightfall, the Cys residues in the four enzymes are reoxidized to their disulfide forms, the enzymes are inactivated, and ATP is not expended in assimilation. Instead, starch synthesized and stored during the daytime is degraded to fuel glycolysis and oxidative phosphorylation at night.
FIGURE 20-37 Light activation of several enzymes of the Calvin cycle. The light activation is mediated by thioredoxin, a small, disulfide-containing protein. In the light, thioredoxin is reduced by electrons moving from photosystem I through ferredoxin (Fd) (blue arrows), then thioredoxin reduces critical disulfide bonds in each of the enzymes sedoheptulose 1,7-bisphosphatase, fructose 1,6-bisphosphatase, ribulose 5-phosphate kinase, and glyceraldehyde 3-phosphate dehydrogenase, activating these enzymes. In the dark, the OSH groups undergo reoxidation to disulfides, inactivating the four enzymes.
Glucose 6-phosphate dehydrogenase, the first enzyme in the oxidative pentose phosphate pathway, is also regulated by this light-driven reduction mechanism, but in the opposite sense. During the day, when photosynthesis produces plenty of NADPH, this enzyme is not needed for NADPH production. Reduction of a critical disulfide bond by electrons from ferredoxin inactivates the enzyme.
SUMMARY 20.4 Carbon-Assimilation Reactions
Photosynthesis in eukaryotes takes place in chloroplasts. In the -assimilating reactions (the Calvin cycle), ATP and NADPH are used to reduce to triose phosphates. These reactions occur in three stages: the fixation reaction itself, catalyzed by rubisco; reduction of the resulting 3-phosphoglycerate to glyceraldehyde 3-phosphate; and regeneration of ribulose 1,5-bisphosphate from triose phosphates.
Stromal enzymes rearrange the carbon skeletons of triose phosphates to generate intermediates of three, four, five, six, and seven carbons, eventually yielding pentose phosphates. The pentose phosphates are converted to ribulose 5-phosphate, which is phosphorylated to ribulose 1,5-bisphosphate to complete the Calvin cycle.
The cost of fixing three into one triose phosphate is nine ATP and six NADPH, which are provided by the light-dependent reactions of photosynthesis.
An antiporter in the inner chloroplast membrane exchanges in the cytosol for 3-phosphoglycerate or dihydroxyacetone phosphate molecules produced by assimilation in the stroma. Oxidation of dihydroxyacetone phosphate in the cytosol generates ATP and NADH, thus moving ATP and reducing equivalents from the chloroplast to the cytosol.
Four enzymes of the Calvin cycle are activated indirectly by light and are inactive in the dark, so that hexose synthesis does not compete with glycolysis — which is required to provide energy in the dark.
 Plants (and other autotrophs) can reduce atmospheric to trioses, then use the trioses as precursors for the synthesis of sucrose and starch, lipids and proteins, and the many other organic components of plant cells (
Plants (and other autotrophs) can reduce atmospheric to trioses, then use the trioses as precursors for the synthesis of sucrose and starch, lipids and proteins, and the many other organic components of plant cells (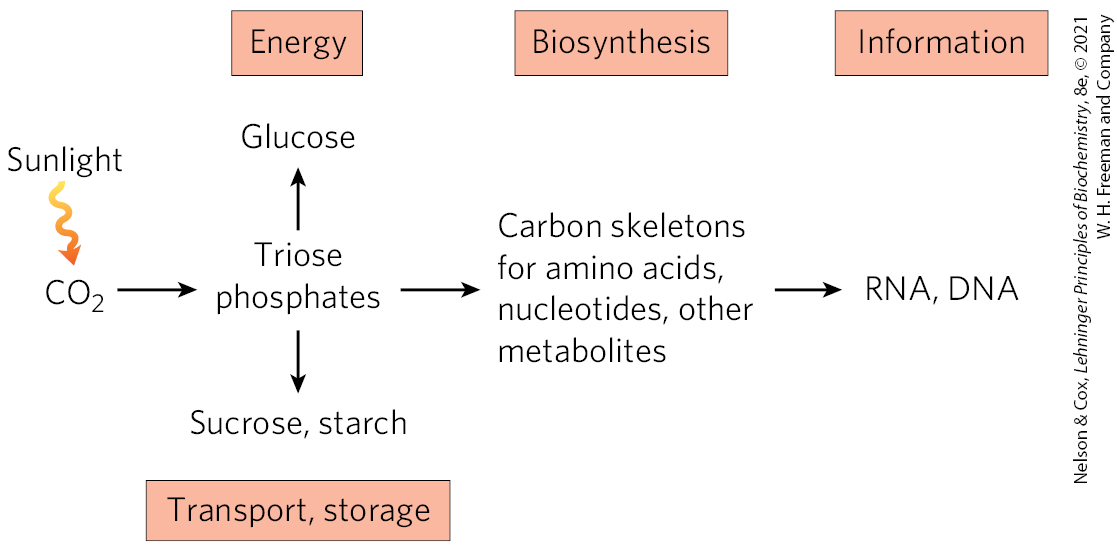

 fructose 1,6-bisphosphatase,
fructose 1,6-bisphosphatase,  sedoheptulose 1,7-bisphosphatase, and
sedoheptulose 1,7-bisphosphatase, and  ribulose 5-phosphate kinase.
ribulose 5-phosphate kinase.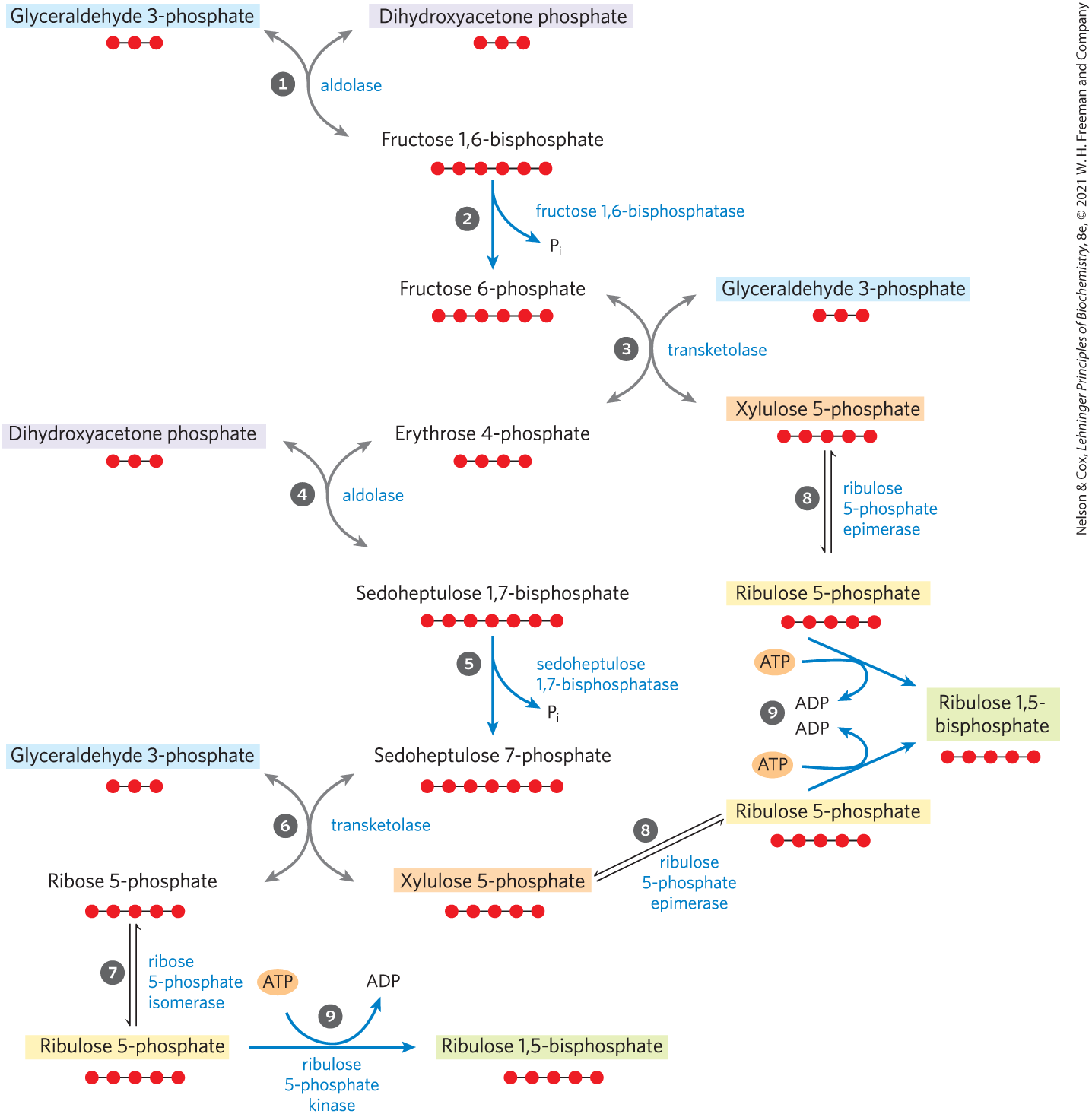
 and
and  ) and transketolase (
) and transketolase ( and
and  ) produce pentose phosphates that are converted to ribulose 1,5-bisphosphate — ribose 5-phosphate by
) produce pentose phosphates that are converted to ribulose 1,5-bisphosphate — ribose 5-phosphate by  ribose 5-phosphate isomerase and xylulose 5-phosphate by
ribose 5-phosphate isomerase and xylulose 5-phosphate by  ribulose 5-phosphate epimerase. Ribulose 5-phosphate is phosphorylated
ribulose 5-phosphate epimerase. Ribulose 5-phosphate is phosphorylated 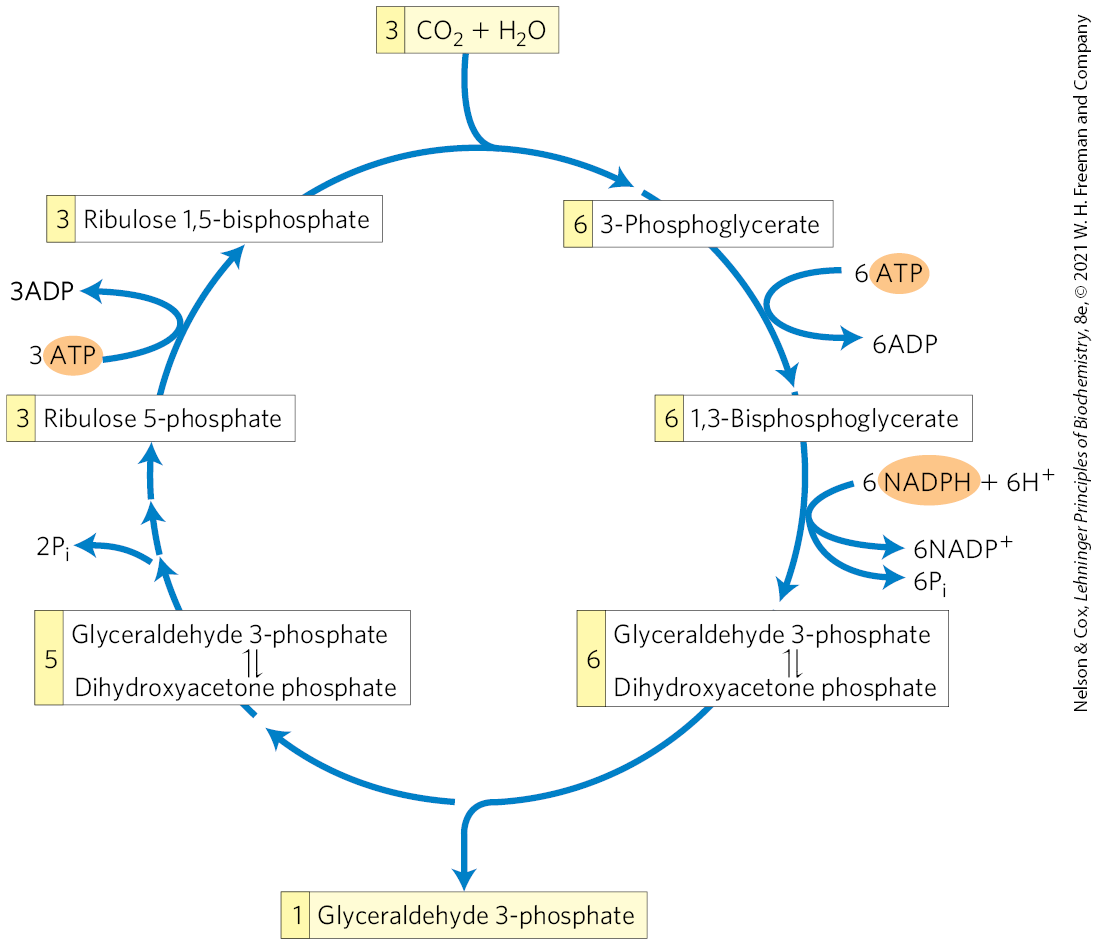

 Photosynthesis in eukaryotes takes place in chloroplasts. In the -assimilating reactions (the Calvin cycle), ATP and NADPH are used to reduce to triose phosphates. These reactions occur in three stages: the fixation reaction itself, catalyzed by rubisco; reduction of the resulting 3-phosphoglycerate to glyceraldehyde 3-phosphate; and regeneration of ribulose 1,5-bisphosphate from triose phosphates.
Photosynthesis in eukaryotes takes place in chloroplasts. In the -assimilating reactions (the Calvin cycle), ATP and NADPH are used to reduce to triose phosphates. These reactions occur in three stages: the fixation reaction itself, catalyzed by rubisco; reduction of the resulting 3-phosphoglycerate to glyceraldehyde 3-phosphate; and regeneration of ribulose 1,5-bisphosphate from triose phosphates.