Studies on a variety of bacteria that carry out photosynthesis have been helpful in understanding the mechanisms of photosynthesis in cyanobacteria, algae and vascular plants. Photosynthetic bacteria have relatively simple phototransduction machinery, with one of two general types of photosystems. Both systems send electrons through a cytochrome complex that pumps protons, producing the electrochemical gradient that drives ATP synthesis.
Photosynthetic Bacteria Have Two Types of Reaction Center
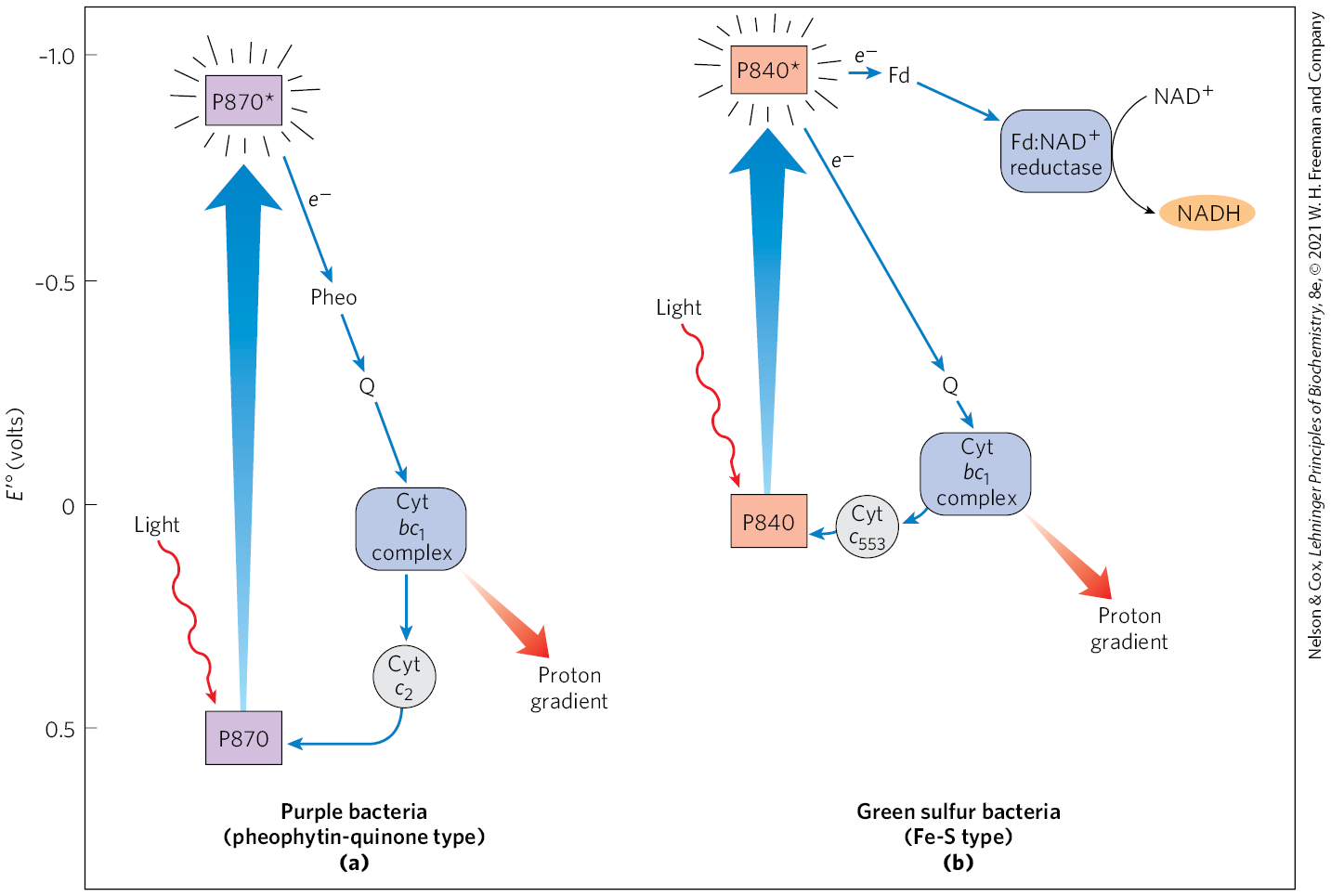
The type II photosystem in purple bacteria consists of three basic modules (Fig. 20-11a): a single P870 reaction center; a cytochrome electron-transfer complex similar to Complex III of the mitochondrial electron-transfer chain; and an ATP synthase, also similar to that of mitochondria. Illumination lifts an electron in the reaction center to its excited state (P870*), from which it passes through pheophytin (chlorophyll a lacking its central ) and a quinone to the cytochrome complex. After passing through the complex, electrons flow through cytochrome back to the reaction center, restoring its preillumination state and completing one cycle. This light-driven cyclic electron transfer provides the energy for proton pumping by the cytochrome complex. Powered by the resulting proton gradient, ATP synthase produces ATP, exactly as in mitochondria.
The type I photosystem in green sulfur bacteria involves the same three modules as in purple bacteria, but the process differs in several respects and includes additional enzymatic reactions (Fig. 20-11b). Excitation by light causes an electron to move from the excited reaction center to the cytochrome complex via a quinone carrier. Electron transfer through this complex powers proton transport and creates the proton-motive force used for ATP synthesis, just as in purple bacteria and in mitochondria. However, in contrast to the cyclic electron transfer path in purple bacteria, some electrons follow a linear electron transfer path from the reaction center to the soluble iron-sulfur protein ferredoxin (see Fig. 19-5), which then passes electrons via ferredoxin: reductase to , producing NADH. The electrons taken from the reaction center to reduce are replaced by the oxidation of to elemental S in the reaction that defines the green sulfur bacteria. This oxidation of by bacteria is chemically analogous to the oxidation of by oxygenic plants. Note that the path of electrons in the purple bacteria is cyclic; the path in the green sulfur bacteria can be either cyclic or linear, leading to and producing NADH.
FIGURE 20-11 Functional Modules of Photosynthetic Machinery in Purple Bacteria and Green Sulfur Bacteria. The position on the vertical scale of each electron carrier reflects its standard reduction potential. (a) In purple bacteria, light energy excites an electron in the reaction center P870. The electron passes through pheophytin (Pheo), a quinone (Q), and the cytochrome complex, then through cytochrome and thus back to the reaction center. Electron transfer through the cytochrome complex causes proton pumping, creating an electrochemical potential that powers ATP synthesis. (b) Green sulfur bacteria have two routes for electrons driven by excitation of P840. A cyclic electron transfer route that goes through a quinone to the cytochrome complex and back to the reaction center via cytochrome causes proton pumping. A linear electron transfer route that goes from the reaction center through the iron-sulfur protein ferredoxin (Fd) reduces to NADH in a reaction catalyzed by ferredoxin: reductase.
In Vascular Plants, Two Reaction Centers Act in Tandem
The photosynthetic apparatus of cyanobacteria, algae, and vascular plants is more complex than the one-center bacterial systems, and it most likely evolved through the combination of two simpler bacterial photosystems. The Z scheme diagram in Figure 20-12 outlines the path of electron flow between the two photosystems and the energy relationships in the light-dependent reactions. (The Z scheme takes its name from the zigzag pattern of the pathways in the diagram.) The thylakoid membranes of chloroplasts have two different kinds of photosystems, each with its own type of photochemical reaction center and set of antenna molecules. The two systems have distinct and complementary functions. Photosystem II (PSII) is a pheophytin-quinone type of system (like the single photosystem of purple bacteria) containing roughly equal amounts of chlorophylls a and b. Excitation of the P680 special pair in its reaction center drives electrons through the cytochrome complex discussed below, with concomitant pumping of protons across the thylakoid membrane and ATP synthesis. Photosystem I (PSI) is structurally and functionally related to the photosynthetic machinery of green sulfur bacteria. It has a P700 reaction center and a high ratio of chlorophyll a to chlorophyll b. The excited P700 passes electrons through a linear chain of carriers to ferredoxin, then to , producing NADPH. An alternative pathway for electrons is cyclic: instead of following the linear path that leads to reduction, electrons pass to plastoquinone (PQ) through a membrane-embedded protein complex, cytochrome (again, with the movement of protons into the chloroplast lumen). The thylakoid membranes of a single spinach chloroplast have many hundreds of each kind of photosystem.
FIGURE 20-12 Integration of photosystems I and II in chloroplasts. This “Z scheme” shows the pathway of linear electron transfer from (lower left) to (far right). The position on the vertical scale of each electron carrier reflects its standard reduction potential. To raise the energy of electrons derived from to the energy level required to reduce to NADPH, each electron must be “lifted” twice (heavy arrows) by photons absorbed in PSII and PSI. One photon is required per electron in each photosystem. After excitation, the high-energy electrons flow “downhill” through the carrier chains as shown. Protons move across the thylakoid membrane during the water-splitting reaction and during electron transfer through the cytochrome complex, producing the proton gradient that is essential to ATP formation. An alternative path of electrons is cyclic electron transfer, in which electrons move from ferredoxin back to the plastoquinone and cytochrome complex, instead of reducing to NADPH. The cyclic pathway produces more ATP and less NADPH than the linear pathway.
These two photosystems in plants act in tandem to catalyze the light-driven movement of electrons from to . The electron carriers include large, integral protein complexes (PSI, PSII, and the proton-pumping complex cytochrome ); quinones that are lipid-soluble and move through the membrane between the protein complexes; and two soluble proteins, plastocyanin (analogous to cytochrome c in mitochondria) and ferredoxin.
To replace the electrons that move from PSII through PSI to is oxidized, producing (Fig. 20-12, bottom left). All -evolving photosynthetic cells — those of plants, algae, and cyanobacteria — contain both PSI and PSII. The Z scheme thus describes the complete route by which electrons flow from to , according to the equation
For every two photons absorbed (one by each photosystem), one electron is transferred from to . To form one molecule of , which requires transfer of four electrons from two to two , a total of eight photons must be absorbed, four by each photosystem.
Having seen the overall process, we’ll now look at how the structure of the photosystems informs our understanding of the electrochemistry.
Photosystem II
PSII is dimeric (Fig. 20-13). Each monomer is a huge complex of 19 proteins, including the accessory proteins CP47 and CP43, and the core complex of P680 reaction-center proteins D1 and D2; 2 chlorophyll-binding proteins; and associated chromophores, including carotenoids, a nonheme iron, and the critically important inorganic cofactor, . Of the proteins in PSII, 16 have transmembrane segments, but 3 are peripheral proteins on the lumenal side that stabilize the cofactor. Surrounding PSII are additional chlorophyll-binding proteins and light-harvesting complexes. When a photon is absorbed by any of these antenna molecules, the resulting exciton moves very rapidly from one to another of the antenna chlorophylls until it reaches the reaction center and excites P680, the special pair of chlorophyll a molecules , to initiate the photochemistry.
FIGURE 20-13 Structure of photosystem II of the cyanobacterium Thermosynechococcus vulcanus. The enormous complex, visualized by x-ray crystallography, is a dimer; each monomer has its own reaction center. Chlorophyll-binding proteins CP43 and CP47 form the core antenna, directly associated with the PSII reaction-center proteins D1 and D2. Each PSII monomer contains 35 chlorophylls, 2 pheophytins, 11 β-carotenes, 2 plastoquinones, and 1 each of b-type cytochrome, c-type cytochrome, and nonheme iron. Water is oxidized to form at the oxygen-evolving center . [Data from PDB ID 3WU2, Y. Umena et al., Nature 473:55, 2011.]
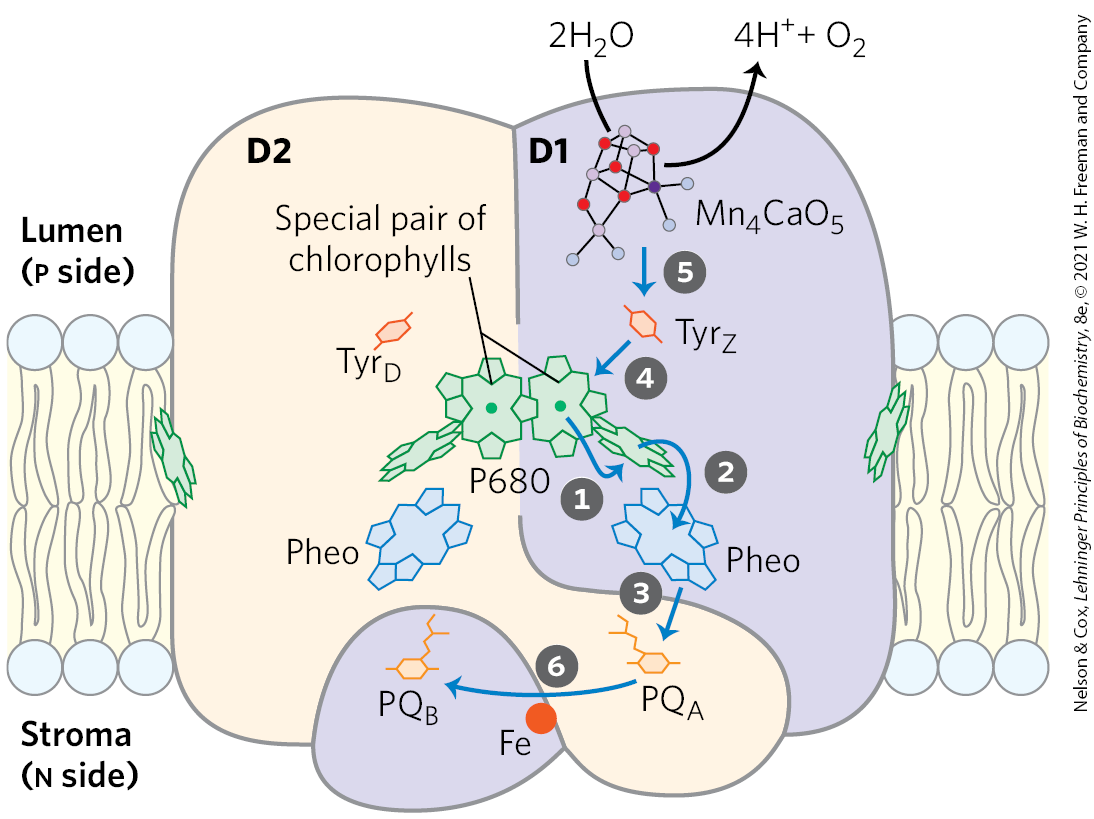
Excitation of P680 in PSII (Fig. 20-14) produces , an excellent electron donor that, within picoseconds, transfers an electron to pheophytin, giving it a negative charge . With the loss of its electron, is transformed into a radical cation, . very rapidly passes its extra electron to a protein-bound plastoquinone, , which in turn passes its electron to another, more loosely bound plastoquinone, . When has acquired two electrons in two such transfers from and two protons from the solvent water, it is in its fully reduced quinol form, . The overall reaction initiated by light in PSII is
(20-1)
Eventually, the electrons in pass through the cytochrome complex (see Fig. 20-12). The electron initially removed from P680 is replaced with an electron obtained from the oxidation of water, as described below.
FIGURE 20-14 Electron transfer through photosystem II of the cyanobacterium Synechococcus elongatus. The monomeric form of the core complex shown here has two major transmembrane proteins, D1 and D2, each with its set of electron carriers. Although the two subunits are nearly symmetrical, electron transfer occurs through only one of the two branches of electron carriers: that on the right (in D1). The arrows show the path of electron transfer from the ion cofactor of the oxygen-evolving complex to plastoquinone . The photochemical events occur in the sequence indicated by the step numbers. The role of the Tyr residues and the detailed structure of the cofactor are discussed below (see Fig. 20-20b).
Photosystem I
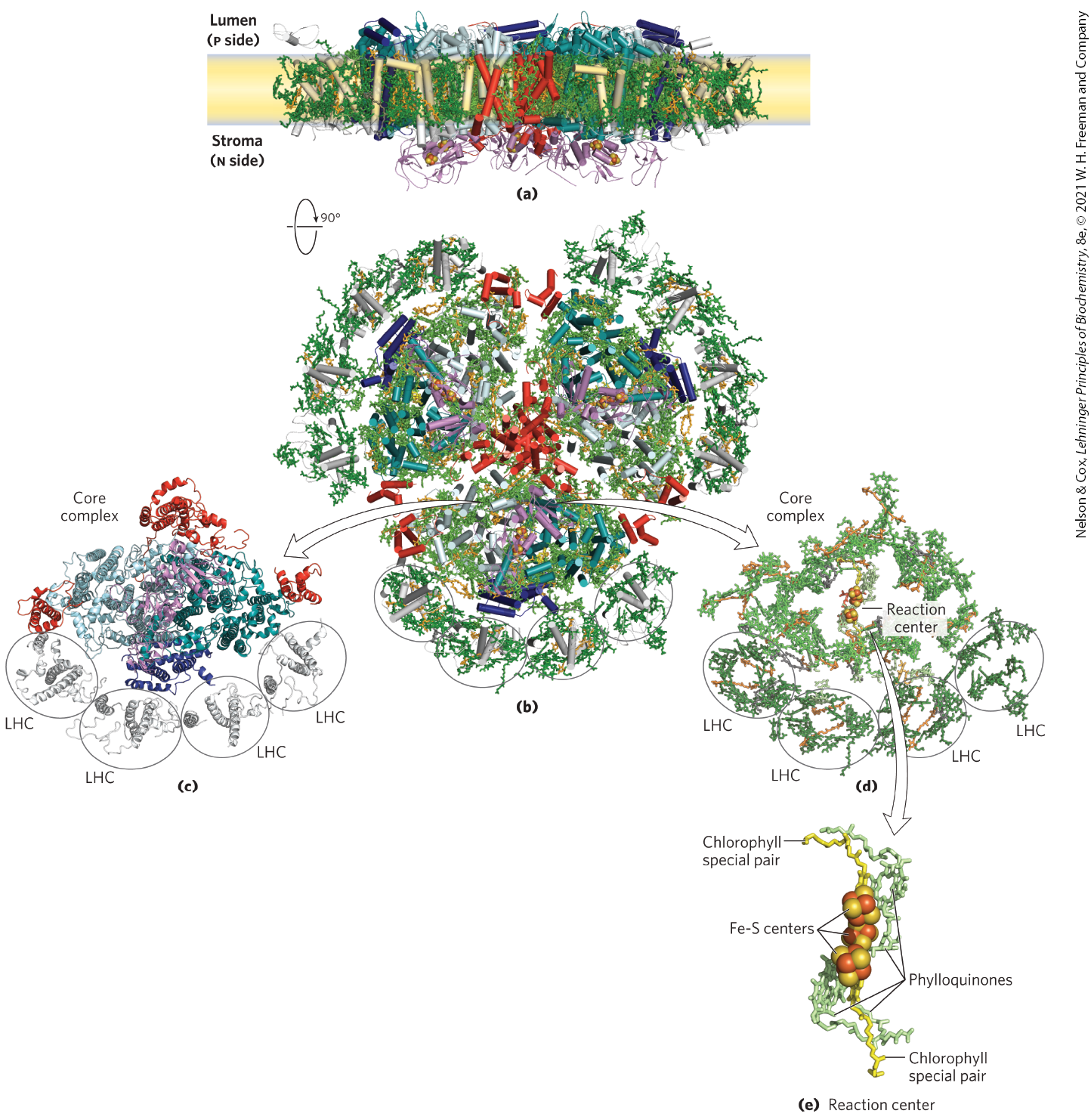
PSI and its antenna molecules are part of a supramolecular complex composed of at least 16 proteins, including 4 chlorophyll-binding proteins arranged around the periphery of the reaction center (Fig. 20-15). The complex also includes 35 carotenoids of several types, three 4Fe-4S clusters, and two phylloquinones.
FIGURE 20-15 Structure of photosystem I in the cyanobacterium Synechococcus elongatus. PSI is a symmetric trimer, viewed here (a) in the plane of the thylakoid membrane and (b) from the stroma (n side of the membrane). (c) One of the three core complexes in PSI, displayed as the protein without its ligands and (d) the ligands alone. Note the four peripheral light-harvesting complexes (LHC) and the many chlorophyll molecules surrounding the reaction center. (e) Close-up view of the reaction center without the surrounding chlorophylls, showing the chlorophyll special pair, phylloquinones, and Fe-S centers. [Data from PDB ID 1JBO, P. Jordan et al., Nature 411:909, 2001; PDB ID 4RKU, Y. Mazor et al.]
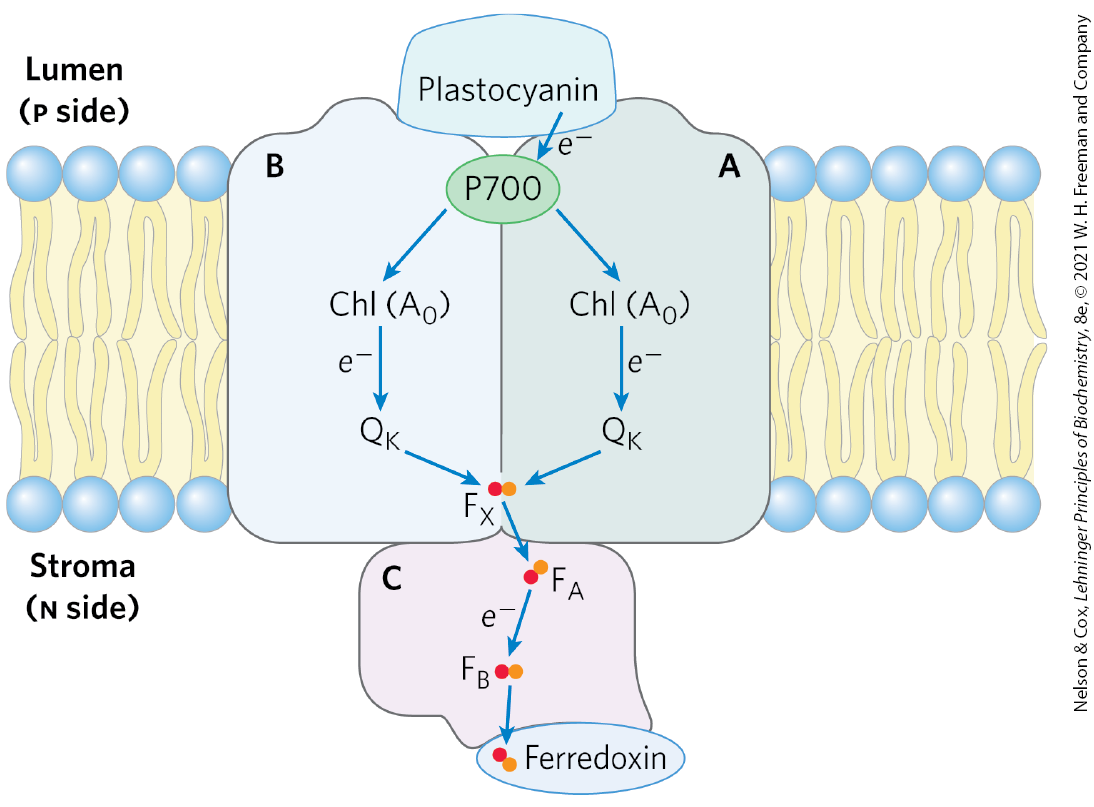
The photochemical events that follow the excitation of PSI at the reaction-center P700 (Fig. 20-16) are formally similar to those occurring in PSII. The excited reaction-center loses an electron to an acceptor, designated (a chlorophyll a molecule, functionally homologous to the pheophytin of PSII), creating and . Again, excitation results in charge separation at the photochemical reaction center. is a strong oxidizing agent, which quickly acquires an electron from plastocyanin, a soluble Cu-containing electron-transfer protein. is an exceptionally strong reducing agent that passes its electron through a chain of carriers that leads to (Fig. 20-12, right side). Phylloquinone accepts the electron and passes it to an iron-sulfur protein through three Fe-S centers in PSI. From here, the electron moves to ferredoxin (Fd). Recall that ferredoxin contains a 2Fe-2S center (see Fig. 19-5) that undergoes one-electron oxidation and reduction reactions. The fourth electron carrier in the chain is the flavoprotein ferredoxin: reductase, which transfers electrons from reduced ferredoxin :
FIGURE 20-16 The path of electrons through PSI. The path of electrons (blue arrows) through PSI, viewed in the plane of the membrane. When the reaction-center P700, the special pair of chlorophylls, is excited by a photon or an exciton, its reduction potential is dramatically reduced, making it a good electron donor. P700 then passes an electron through a nearby chlorophyll (referred to as ) to phylloquinone . Reduced is reoxidized as it passes two electrons, one at a time, to an Fe-S center near the n side of the membrane. From , electrons move through two more Fe-S centers to ferredoxin in the stroma. Ferredoxin then donates electrons to (not shown), reducing it to NADPH, one of the forms in which the energy of photons is trapped in chloroplasts.
The Cytochrome Complex Links Photosystems II and I, Conserving the Energy of Electron Transfer
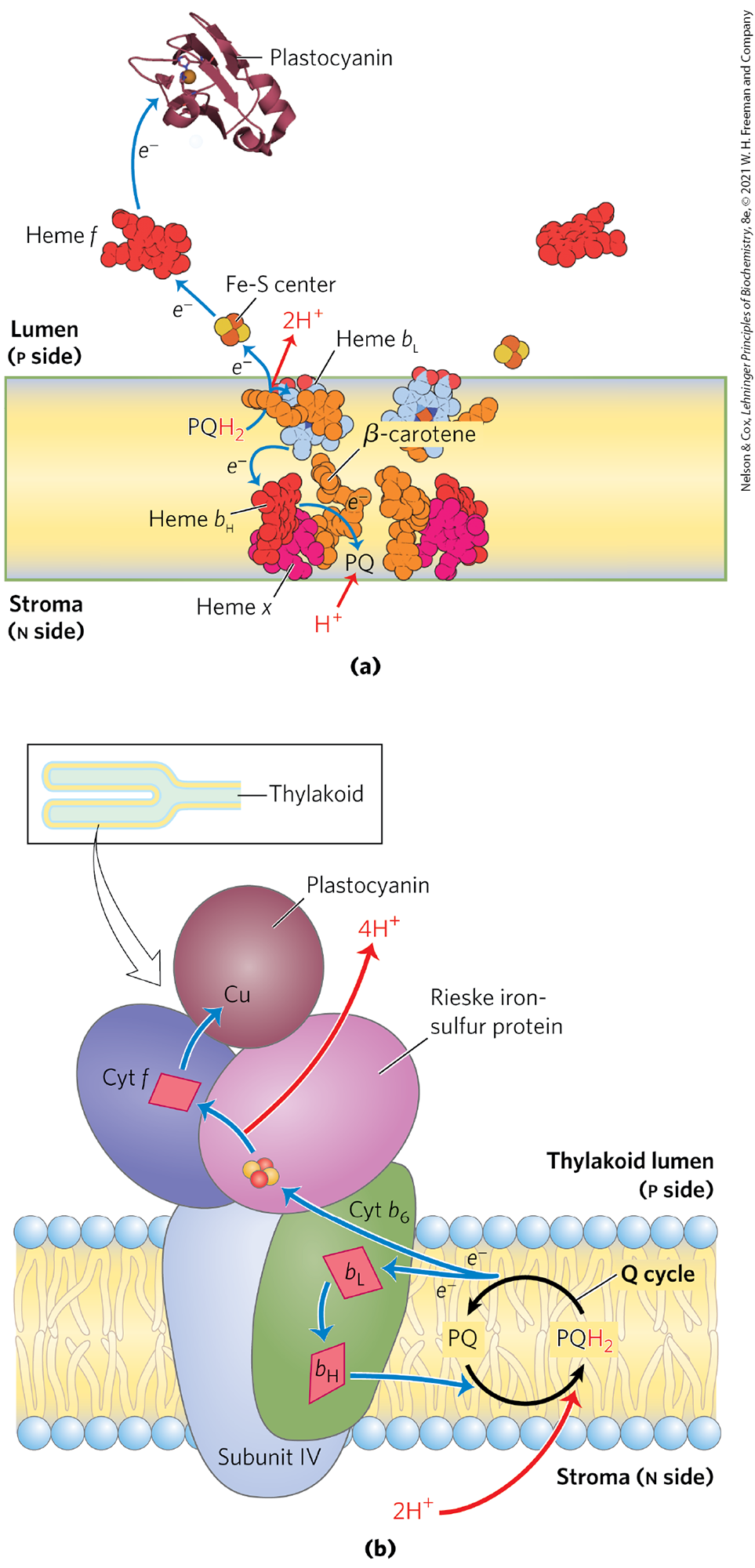
Electrons temporarily held in plastoquinol as a result of the excitation of P680 in PSII are carried to P700 of PSI via the cytochrome complex and the soluble protein plastocyanin (see Fig. 20-12, center). With a structure and role analogous with that of Complex III in mitochondria, the cytochrome complex (Fig. 20-17) contains a b-type cytochrome with two heme groups (designated and ), a Rieske iron-sulfur protein , and cytochrome f (named for the Latin frons, “leaf”). Electrons flow through the cytochrome complex from to cytochrome f, then to plastocyanin, and finally to , thereby reducing it.
FIGURE 20-17 Electron and proton flow through the cytochrome complex. (a) In addition to the hemes of cytochrome b (heme and ; also called heme and , respectively, because of their proximity to the n and p sides of the bilayer) and cytochrome f (heme f), there is a fourth heme (heme x) near heme ; also present is a β-carotene of unknown function. Two sites bind plastoquinone: the site near the p side of the bilayer, and the PQ site near the n side. The Fe-S center of the Rieske protein lies just outside the bilayer on the p side, and the heme f site is on a protein domain that extends well into the thylakoid lumen. The electron path is shown for just one of the monomers, but both sets of carriers in the dimer carry electrons to plastocyanin. (b) Plastoquinol , formed in PSII, is oxidized by the cytochrome complex in a series of steps like those of the Q cycle in Complex III of mitochondria (see Fig. 19-11). One electron from passes to the Fe-S center of the Rieske protein, the other to heme of cytochrome . The net effect is passage of electrons from to the soluble protein plastocyanin, which carries them to PSI. [Data from PDB ID 1VF5, G. Kurisu et al., Science 302:1009, 2003; PDB ID 2Q5B, Y. S. Bukhman-DeRuyter et al.]
Like Complex III of mitochondria, cytochrome conveys electrons from a reduced quinone — a mobile, lipid-soluble carrier of two electrons (Q in mitochondria, in chloroplasts; P for plastoquinone) — to a water-soluble protein that carries one electron (cytochrome c in mitochondria, plastocyanin in chloroplasts) (Fig 20-17a). As in mitochondria, the function of this complex involves a Q cycle (Fig. 20-17b; see Fig. 19-11) in which electrons pass, one at a time, from to cytochrome . This cycle results in the pumping of protons across the membrane, from the stromal compartment to the thylakoid lumen. Up to four protons enter the lumen for each pair of electrons that passes through the cytochrome complex. The result is production of a proton gradient across the thylakoid membrane as electrons pass from PSII to PSI. Because the volume of the flattened thylakoid lumen is small, the influx of a small number of protons has a relatively large effect on lumenal pH. The measured difference in pH between the stroma (pH 8) and the thylakoid lumen (pH 5) represents a 1,000-fold difference in proton concentration — a powerful driving force for ATP synthesis.
Cyclic Electron Transfer Allows Variation in the Ratio of ATP/NADPH Synthesized
Cyclic electron flow between PSI and cytochrome increases the production of ATP relative to NADPH. The linear path of electrons from water, through PSII, cytochrome , and PSI to produces both a proton gradient, which is used to drive ATP synthesis, and NADPH, which is used in reductive biosynthetic processes (see Fig. 20-12). Some fraction of electrons passing from to ferredoxin do not continue to , but cycle back through plastoquinone and the cytochrome complex to plastocyanin. Plastocyanin then donates electrons to P700. In this way, electrons are repeatedly recycled through the cytochrome complex and the reaction center of PSI, each electron propelled around the cycle by the energy of one photon. Cyclic electron flow is not accompanied by net formation of NADPH or evolution of . However, it is accompanied by proton pumping by the cytochrome complex and by phosphorylation of ADP to ATP, referred to as cyclic photophosphorylation. The overall equation for cyclic electron flow and photophosphorylation is simply
By regulating the partitioning of electrons between reduction and cyclic photophosphorylation, a plant adjusts the ratio of ATP to NADPH produced in the light-dependent reactions to match its needs for these products in the -assimilation reactions and other biosynthetic processes. As we shall see in Section 20.4, the -assimilation reactions require ATP and NADPH in the ratio 3:2. This regulation of electron-transfer pathways is part of a short-term adaptation to changes in light color (wavelength) and quantity (intensity).
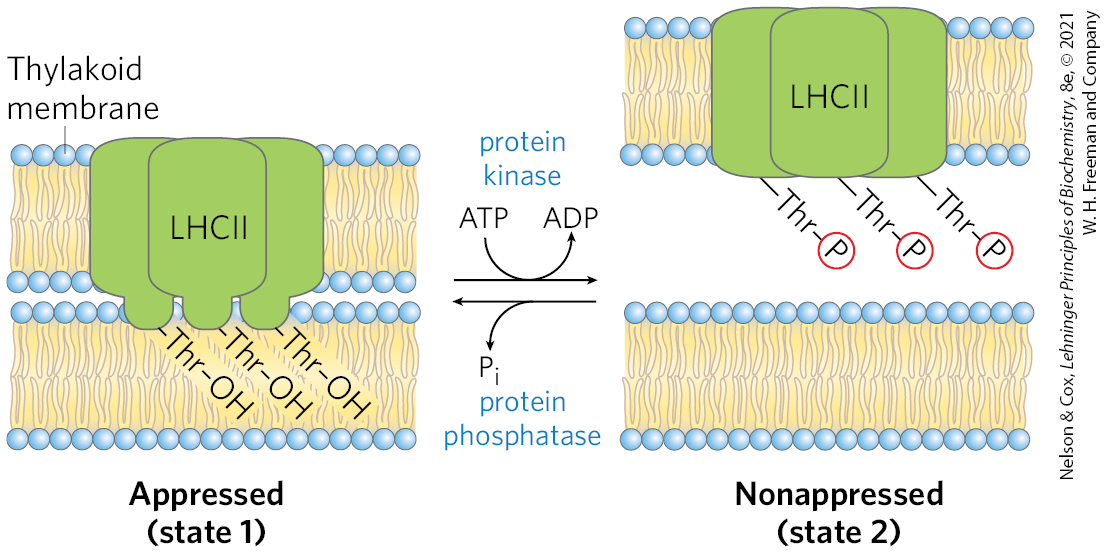
State Transitions Change the Distribution of LHCII between the Two Photosystems
Photosynthetic organisms are exposed to light of highly variable intensity and wavelength in the course of a day or a season, and, although they can alter their growth patterns somewhat, they cannot uproot themselves and move to optimize their light exposure. Instead, cellular mechanisms have evolved that allow plants to accommodate changing light conditions. The energy needed to excite PSI (P700) is less (light of longer wavelength, lower energy) than the energy needed to excite PSII (P680). If PSI and PSII were physically contiguous, excitons originating in the antenna system of PSII would migrate to the reaction center of PSI, leaving PSII chronically underexcited and thus interfering with the operation of the two-center system. This imbalance in the supply of excitons is prevented by physically separating the two photosystems in the thylakoid membrane (Fig. 20-18). PSII is located almost exclusively in the tightly appressed membrane stacks of granal thylakoids; its associated light-harvesting complex (LHCII) mediates the tight association of adjacent membranes in the grana. PSI and the ATP synthase complex are located almost exclusively in the nonappressed membranes of the stromal thylakoids, where they have access to the contents of the stroma, including ADP and . The cytochrome complex is present primarily in the granal thylakoids.
FIGURE 20-18 Localization of PSI and PSII in thylakoid membranes. (a) Structures of the complexes and soluble proteins of the photosynthetic apparatus of a vascular plant or alga, drawn to the same scale. The bovine ATP synthase is shown. (b) Light-harvesting complex LHCII and ATP synthase are located both in appressed regions of the thylakoid membrane (granal thylakoids, in which several membranes are in contact) and in nonappressed regions (stromal thylakoids), and have ready access to ADP and in the stroma. PSII is present almost exclusively in the appressed granal regions, and PSI almost exclusively in nonappressed stromal regions. LHCII is the “adhesive” that holds appressed thylakoid membranes together (see Fig. 20-19). [(a) Data from PSII: PDB ID 3WU2, Y. Umena et al., Nature 473:55, 2011; cyt complex: PDB ID 2E74, E. Yamashita et al., J. Mol. Biol. 370:39, 2007; plastocyanin: PDB ID 1AG6, Y. Xue et al., Protein Sci. 7:2099, 1998; PSI: PDB ID 4RKU, Y. Mazor et al.; ferredoxin: PDB ID 1A70, C. Binda et al., Acta Crystallogr. D Biol. Crystallogr. 54:1353, 1998; ferredoxin:NADP reductase: PDB ID 1QG0, Z. Deng et al., Nat. Struct. Biol. 6:847, 1999; ATP synthase: PDB ID 5ARA, A. Zhou et al., eLife 4:e10180, 2015.]
The association of LHCII with PSI and PSII depends on light intensity and wavelength, which can change in the short term and lead to state transitions in the chloroplast. In state 1, LHCII, PSII, and PSI are poised to maximize the capture of light energy. A critical Thr residue in LHCII is unphosphorylated, and LHCII associates with PSII. Under conditions of intense or blue light, which favor absorption by PSII, that photosystem reduces plastoquinone to plastoquinol faster than PSI can oxidize it. The resulting accumulation of activates a protein kinase that triggers the transition to state 2 by phosphorylating a Thr residue on LHCII (Fig. 20-19). Phosphorylation weakens the interaction of LHCII with the appressed membrane and with PSII; some LHCII dissociates and moves to the stromal thylakoids. Here it captures photons (excitons) for PSI, speeding the oxidation of and reversing the imbalance between electron flow in PSI and PSII. In less intense light (in the shade, with more red light), PSI oxidizes faster than PSII can make it, and the resulting increase in [PQ] triggers dephosphorylation of LHCII, reversing the effect of phosphorylation. The state transition in LHCII localization and the transition from cyclic to linear electron transfer are coordinately regulated: the path of electrons is primarily linear in state 1 and primarily cyclic in state 2.
FIGURE 20-19 Electron transfer in PSI and PSII is balanced through state transitions. In granal thylakoids, a hydrophobic domain of LHCII in one membrane inserts into the neighboring membrane and closely appresses the two (state 1). Accumulation of plastoquinol (not shown) stimulates a protein kinase that phosphorylates a Thr residue in the hydrophobic domain of LHCII, which reduces its affinity for the neighboring membrane and converts appressed granal thylakoids to nonappressed stromal thylakoids (state 2). A specific protein phosphatase reverses this regulatory phosphorylation when the ratio increases.
When light is so intense that the combined activity of PSII and PSI cannot synthesize ATP and NADPH fast enough to keep up with the supply of photons, carotenoids in LHCII absorb the excitons and very rapidly quench the excited chlorophyll before it can create damaging reactive oxygen species (ROS). The trigger for switching from an efficient light-harvesting state to an energy-dissipating state is the lowering of pH in the lumenal space, but the detailed mechanism for this transition is not yet known.
Water Is Split at the Oxygen-Evolving Center
The ultimate source of the electrons passed to NADPH in plant (oxygenic) photosynthesis is water. Having given up an electron to pheophytin, (of PSII) must acquire an electron to return to its ground state in preparation for capture of another photon. In principle, the required electron might come from any number of organic or inorganic compounds. Photosynthetic bacteria use a variety of electron donors for this purpose — acetate, succinate, malate, or sulfide — depending on what is available in a particular ecological niche. About 2.5 billion years ago, evolution of primitive photosynthetic bacteria (progenitors of the modern cyanobacteria) produced a photosystem capable of taking electrons from a donor that is always available: water. Two water molecules are oxidized, yielding four electrons, four protons, and molecular oxygen:
A single photon of visible light does not have enough energy to break the bonds in water; four photons are required in this photolytic cleavage reaction.
The four electrons abstracted from water do not pass directly to , which can accept only one electron at a time. Instead, a remarkable molecular device, the oxygen-evolving center, passes four electrons one at a time to (Fig. 20-20a). The immediate electron donor to is a Tyr residue (sometimes designated Z or ) in subunit D1 of the PSII reaction center. The Tyr residue loses both a proton and an electron, generating the electrically neutral Tyr free radical, :
(20-2)
The Tyr radical regains its missing electron and proton by oxidizing a cofactor of four manganese ions and one calcium ion in the oxygen-evolving center. With each single-electron transfer, the cofactor becomes more oxidized; four single-electron transfers, each corresponding to the absorption of one photon, produce a charge of on the cofactor (Fig. 20-20a):
(20-3)
In this state, the cofactor can take four electrons from a pair of water molecules, releasing four and :
(20-4)
Because the four protons produced in this reaction are released into the thylakoid lumen, the oxygen-evolving center acts as a proton pump, driven by electron transfer.
We saw in Equation 20-1 that the overall reaction initiated by light in PSII is
FIGURE 20-20 Water-splitting activity of the oxygen-evolving center. (a) The process that produces a four-electron oxidizing agent — a multinuclear center with four Mn ions, one Ca ion, and five oxygen atoms — in the oxygen-evolving center of PSII. The sequential absorption of four photons (excitons), each absorption causing the loss of one electron from the cofactor, produces an oxidizing agent that can remove four electrons from two molecules of water, producing . The electrons lost from the cofactor pass one at a time to an oxidized Tyr residue in a PSII protein, then to . (b) The chair-shaped metallic center of the oxygen-evolving center. , known to participate in the oxidation of water, is seen hydrogen-bonded to a network of water molecules, including several directly in contact with the cofactor. This is the site of one of the most important reactions in the biosphere. [(b) Data from PDB ID 3WU2, Y. Umena et al., Nature 473:55, 2011.]
The oxygen-evolving cofactor takes the shape of a chair (Fig. 20-20b). The seat and legs of the chair are made up of three Mn ions, one Ca ion, and four O atoms; the fourth Mn and another O form the back of the chair. Four water molecules are also seen in the crystal structure, two associated with one of the Mn ions, the other two with the Ca ion. It may be one (or more) of these water molecules that undergoes oxidation to produce . This metal cofactor is associated with several peripheral membrane proteins on the lumenal side of the thylakoid membrane that are believed to stabilize the cofactor. The Tyr residue designated Z, through which electrons move between water and the PSII reaction center, is connected with a network of hydrogen-bonded water molecules that includes the four associated with the cofactor. The detailed mechanism of water oxidation by the cofactor is not known but is under intense investigation. The reaction is central to life on Earth and may involve novel bioinorganic chemistry. Determination of the structure of the polymetallic center has inspired several reasonable and testable hypotheses. Stay tuned.
SUMMARY 20.2 Photochemical Reaction Centers
Bacteria have a single photochemical reaction center. Purple bacteria have a type II photosystem where electrons from an excited special pair of chlorophyll molecules flow through pheophytin, quinones, and a proton-pumping cytochrome complex, back to the special pair of chlorophylls. Green sulfur bacteria have a type I photosystem that can send electrons through a similar cyclic path or through a linear path that reduces to NADH.
In cyanobacteria, algae, and plants, two different reaction centers are arranged in tandem. In the reaction center of PSII, when the special pair of chlorophylls (P680) is excited by light, it passes electrons to plastoquinone, and the electrons lost from P680 are replaced by electrons from . PSI passes electrons from the excited special pair in its reaction center through a series of carriers to ferredoxin, which then reduces to NADPH.
Electron flow from either photosystem through the cytochrome complex drives protons across the thylakoid membrane, creating a proton-motive force that provides the energy for ATP synthesis by an ATP synthase.
Linear electron transfer through the photosystems produces NADPH and ATP. Cyclic electron transfer produces only ATP and allows variability in the proportions of NADPH and ATP formed.
The distribution of PSI and PSII between the granal and stromal thylakoids can change and is indirectly controlled by light intensity, optimizing the distribution of excitons between PSI and PSII for efficient energy capture.
The oxygen-evolving center, which contains a cofactor, uses energy from light to split water, producing . For each formed at the oxygen-evolving center, four protons are pumped into the thylakoid lumen, contributing to the proton motive force.
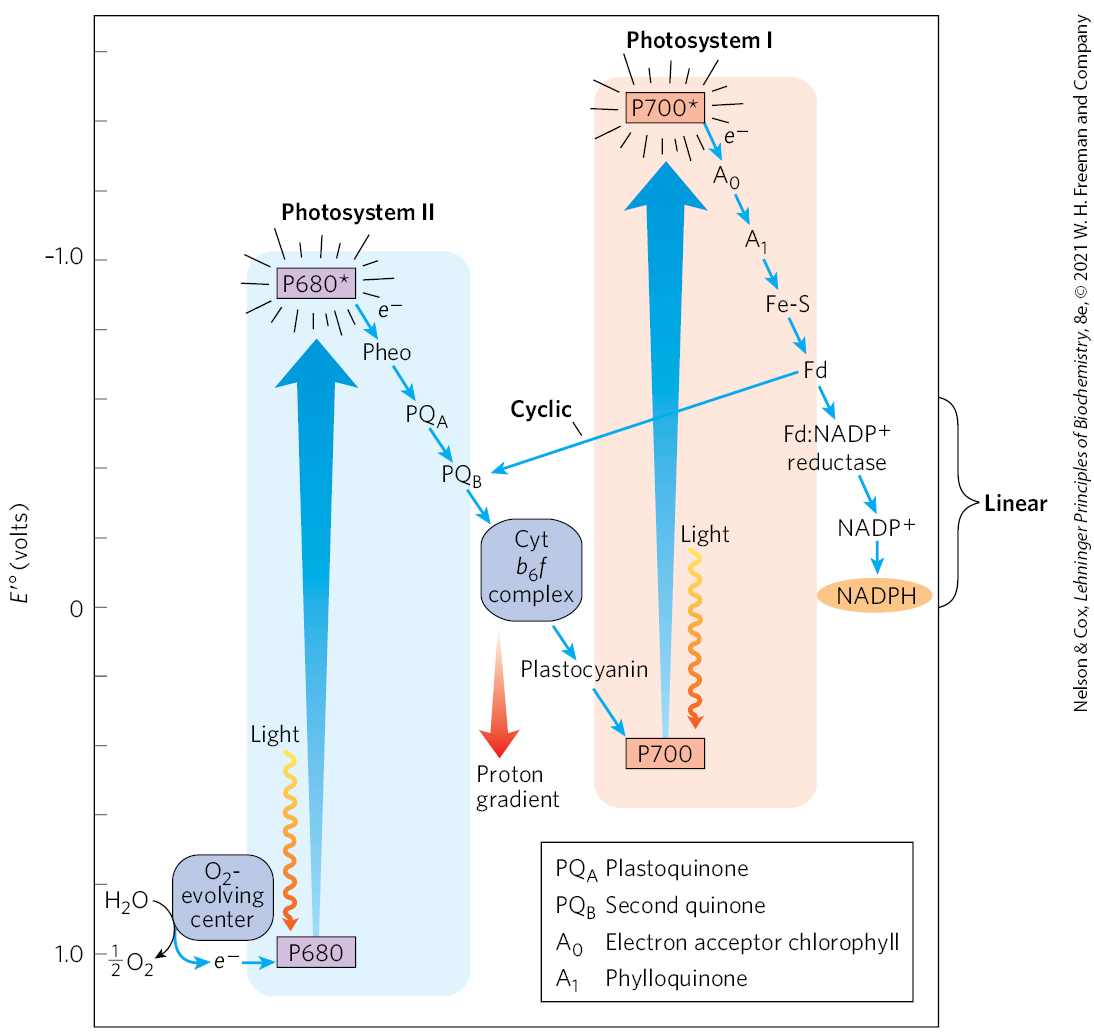
 The Z scheme thus describes the complete route by which electrons flow from to , according to the equation
The Z scheme thus describes the complete route by which electrons flow from to , according to the equation When a photon is absorbed by any of these antenna molecules, the resulting exciton moves very rapidly from one to another of the antenna chlorophylls until it reaches the reaction center and excites P680, the special pair of chlorophyll a molecules , to initiate the photochemistry.
When a photon is absorbed by any of these antenna molecules, the resulting exciton moves very rapidly from one to another of the antenna chlorophylls until it reaches the reaction center and excites P680, the special pair of chlorophyll a molecules , to initiate the photochemistry.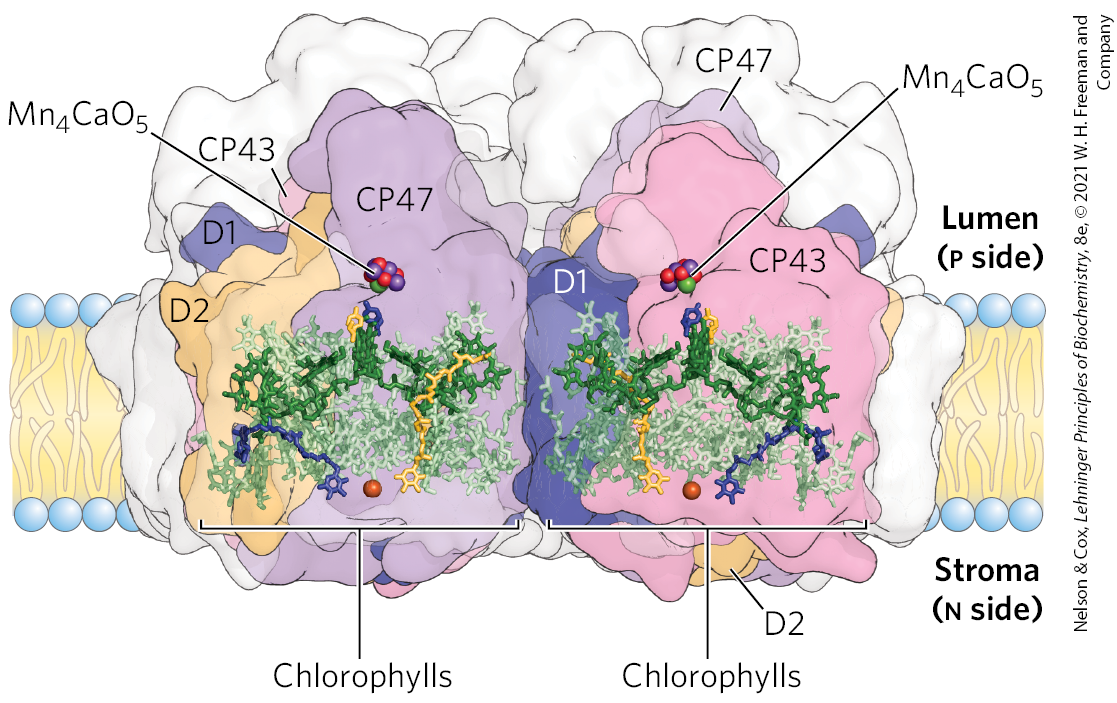
 By regulating the partitioning of electrons between reduction and cyclic photophosphorylation, a plant adjusts the ratio of ATP to NADPH produced in the light-dependent reactions to match its needs for these products in the -assimilation reactions and other biosynthetic processes. As we shall see in
By regulating the partitioning of electrons between reduction and cyclic photophosphorylation, a plant adjusts the ratio of ATP to NADPH produced in the light-dependent reactions to match its needs for these products in the -assimilation reactions and other biosynthetic processes. As we shall see in 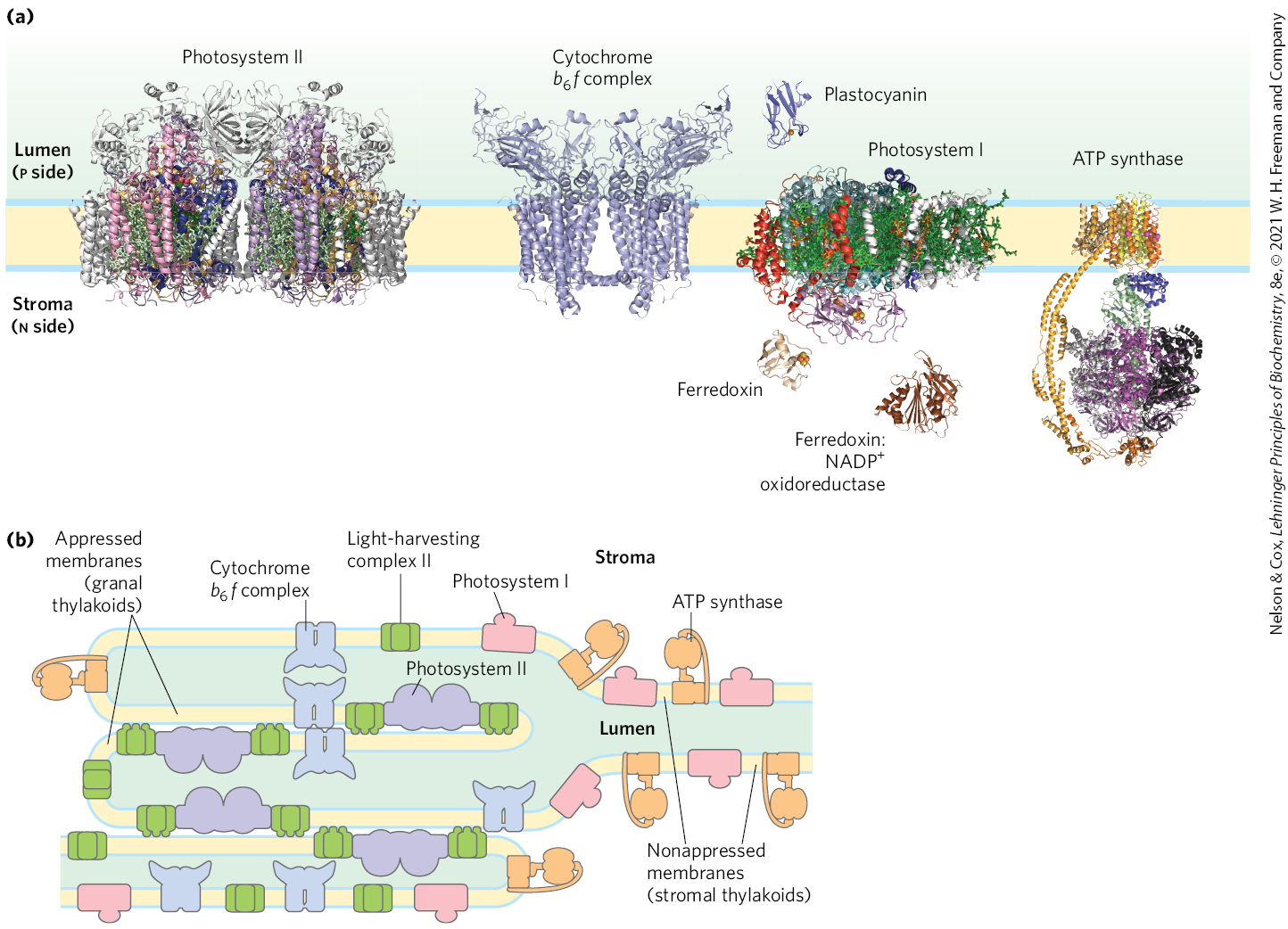
 Because the four protons produced in this reaction are released into the thylakoid lumen, the oxygen-evolving center acts as a proton pump, driven by electron transfer.
Because the four protons produced in this reaction are released into the thylakoid lumen, the oxygen-evolving center acts as a proton pump, driven by electron transfer.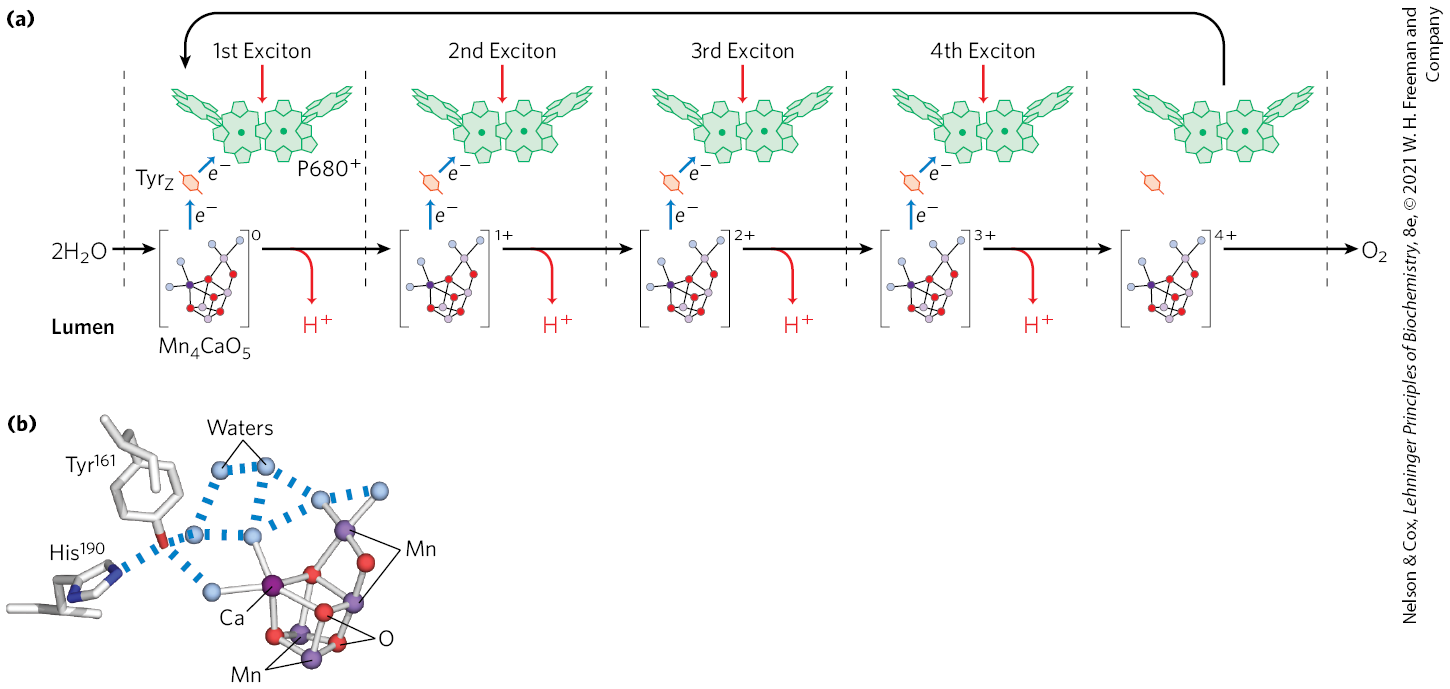
 Bacteria have a single photochemical reaction center. Purple bacteria have a type II photosystem where electrons from an excited special pair of chlorophyll molecules flow through pheophytin, quinones, and a proton-pumping cytochrome complex, back to the special pair of chlorophylls. Green sulfur bacteria have a type I photosystem that can send electrons through a similar cyclic path or through a linear path that reduces to NADH.
Bacteria have a single photochemical reaction center. Purple bacteria have a type II photosystem where electrons from an excited special pair of chlorophyll molecules flow through pheophytin, quinones, and a proton-pumping cytochrome complex, back to the special pair of chlorophylls. Green sulfur bacteria have a type I photosystem that can send electrons through a similar cyclic path or through a linear path that reduces to NADH.