In addition to their role as the building blocks of proteins, amino acids are precursors of many specialized biomolecules, including hormones, coenzymes, nucleotides, alkaloids, cell wall polymers, porphyrins, antibiotics, pigments, and neurotransmitters. We describe here the pathways to a number of these amino acid derivatives.
Glycine Is a Precursor of Porphyrins
The biosynthesis of porphyrins, for which glycine is a major precursor, is our first example because of the central importance of the porphyrin nucleus in heme proteins such as hemoglobin and the cytochromes. The porphyrins are constructed from four molecules of the monopyrrole derivative porphobilinogen, which itself is derived from two molecules of δ-aminolevulinate. There are two major pathways to δ-aminolevulinate. In higher eukaryotes (Fig. 22-25a), glycine reacts with succinyl-CoA in the first step to yield α-amino-β-ketoadipate, which is then decarboxylated to δ-aminolevulinate. In plants, algae, and most bacteria, δ-aminolevulinate is formed from glutamate (Fig. 22-25b). The glutamate is first esterified to ; reduction by NADPH converts the glutamate to glutamate 1-semialdehyde, which is cleaved from the tRNA. An aminotransferase converts the glutamate 1-semialdehyde to δ-aminolevulinate.
FIGURE 22-25 Biosynthesis of δ-aminolevulinate. (a) In most animals, including mammals, δ-aminolevulinate is synthesized from glycine and succinyl-CoA. The atoms furnished by glycine are shown in red. (b) In bacteria and plants, the precursor of δ-aminolevulinate is glutamate.
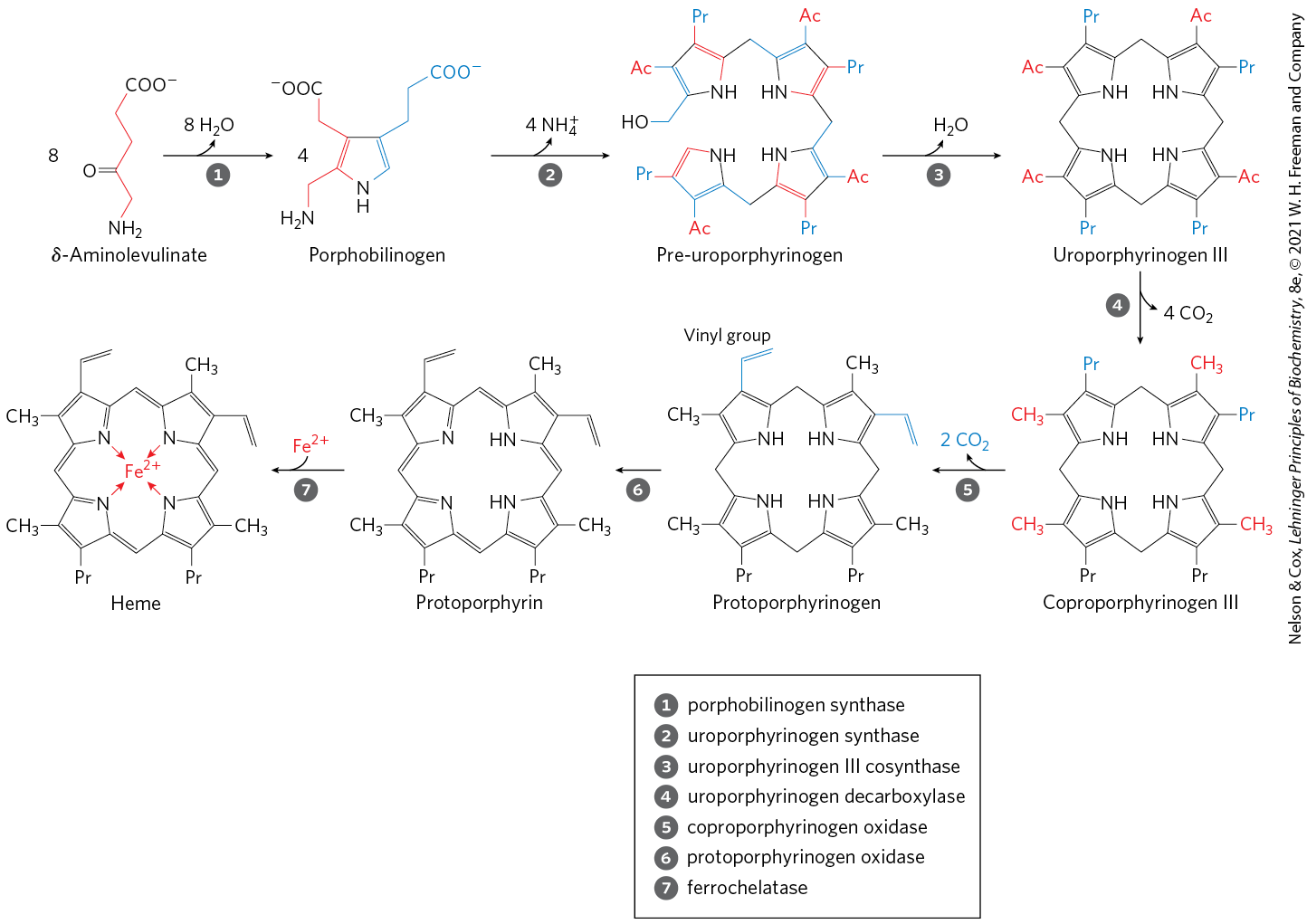
In all organisms, two molecules of δ-aminolevulinate condense to form porphobilinogen and, through a series of complex enzymatic reactions, four molecules of porphobilinogen come together to form protoporphyrin (Fig. 22-26). The iron atom is incorporated after the protoporphyrin has been assembled, in a step catalyzed by ferrochelatase. Porphyrin biosynthesis is regulated in higher eukaryotes by heme, which serves as a feedback inhibitor of early steps in the synthetic pathway. Genetic defects in the biosynthesis of porphyrins can lead to the accumulation of pathway intermediates, causing a variety of human diseases known collectively as porphyrias (Box 22-2).
FIGURE 22-26 Biosynthesis of heme from δ-aminolevulinate. Ac represents acetyl ; Pr, propionyl .
Heme Degradation Has Multiple Functions
The iron-porphyrin (heme) group of hemoglobin, released from dying erythrocytes in the spleen, is degraded to yield free and, ultimately, bilirubin. The pathway also contributes the pigment present in mixtures of the bile salts derived from cholesterol.
The first step in the two-step pathway to bilirubin, catalyzed by heme oxygenase, converts heme to biliverdin, a linear (open) tetrapyrrole derivative (Fig. 22-27 on p. 820). The other products of the reaction are free and CO. The is quickly bound by ferritin. Carbon monoxide is a poison that binds to hemoglobin (see Box 5-1), and the production of CO by heme oxygenase ensures that, even in the absence of environmental exposure, about 1% of an individual’s heme is complexed with CO.
FIGURE 22-27 Bilirubin and its breakdown products. M represents methyl; V, vinyl; Pr, propionyl; E, ethyl. For ease of comparison, these structures are shown in linear form, rather than in their correct stereochemical conformations.
Biliverdin is converted to bilirubin in the second step, catalyzed by biliverdin reductase. You can monitor the reactions in the breakdown of heme (Fig. 22-27) colorimetrically in a familiar in situ experiment. When you are bruised, the black and/or purple color results from hemoglobin released from damaged erythrocytes. Over time, the color changes to the green of biliverdin, and then to the yellow of bilirubin. Bilirubin is largely insoluble, and it travels in the bloodstream as one of many metabolites, fatty acids and others, complexed with serum albumin (see Fig. 17-3). In the liver, bilirubin is transformed to the bile pigment bilirubin diglucuronide. This product is sufficiently water-soluble to be secreted with other components of bile into the small intestine, where microbial enzymes convert it to several products, predominantly urobilinogen. Some urobilinogen is reabsorbed into the blood and transported to the kidney, where it is converted to urobilin, the compound that gives urine its yellow color. Urobilinogen remaining in the intestine is converted (in another microbe-dependent reaction) to stercobilin, which imparts the red-brown color to feces.
Impaired liver function or blocked bile secretion causes bilirubin to leak from the liver into the blood, resulting in a yellowing of the skin and eyeballs, a condition called jaundice. In cases of jaundice, determination of the concentration of bilirubin in the blood may be useful in the diagnosis of underlying liver disease. Newborn infants sometimes develop jaundice because they have not yet produced enough glucuronyl bilirubin transferase to process their bilirubin. A traditional treatment to reduce excess bilirubin, exposure to a fluorescent lamp, causes a photochemical conversion of bilirubin to compounds that are more soluble and easily excreted.
These pathways of heme breakdown play significant roles in protecting cells from oxidative damage and in regulating certain cellular functions. The CO produced by heme oxygenase is toxic at high concentrations, but at the very low concentrations generated during heme degradation it seems to have some regulatory and/or signaling functions. It acts as a vasodilator, much the same as (but less potent than) nitric oxide (discussed below). Low levels of CO also have some regulatory effects on neurotransmission. Bilirubin is the most abundant antioxidant in mammalian tissues and is responsible for most of the antioxidant activity in serum. Its protective effects seem to be especially important in the developing brain of newborn infants. The cell toxicity associated with jaundice may be due to bilirubin levels in excess of the serum albumin needed to solubilize it.
Given these varied roles of heme degradation products, the degradative pathway is subject to regulation, mainly at the first step. Humans have at least three isozymes of heme oxygenase (HO). HO-1 is highly regulated; the expression of its gene is induced by a wide range of stress conditions, including shear stress, uncontrolled angiogenesis (development of blood vessels), hypoxia, hyperoxia, heat shock, exposure to ultraviolet light, hydrogen peroxide, and many other metabolic insults. HO-2 is found mainly in the brain and the testes, where it is continuously expressed. The third isozyme, HO-3, is not catalytically active, but may play a role in oxygen sensing.
Amino Acids Are Precursors of Creatine and Glutathione
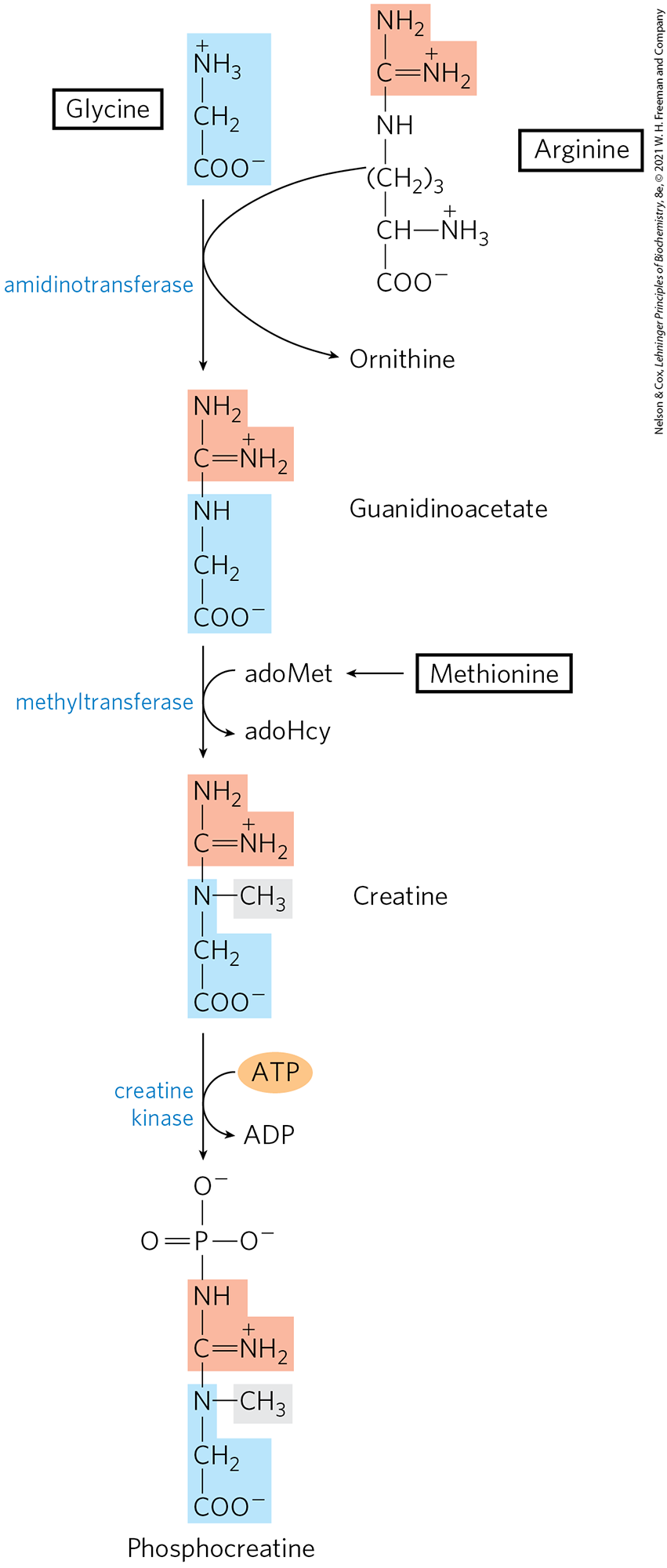
Phosphocreatine, derived from creatine, is an important energy buffer in skeletal muscle (see Box 23-1). Creatine is synthesized from glycine and arginine (Fig. 22-28); methionine, in the form of S-adenosylmethionine, acts as methyl group donor.
FIGURE 22-28 Biosynthesis of creatine and phosphocreatine. Creatine is made from three amino acids: glycine, arginine, and methionine. This pathway shows the versatility of amino acids as precursors of other nitrogenous biomolecules.
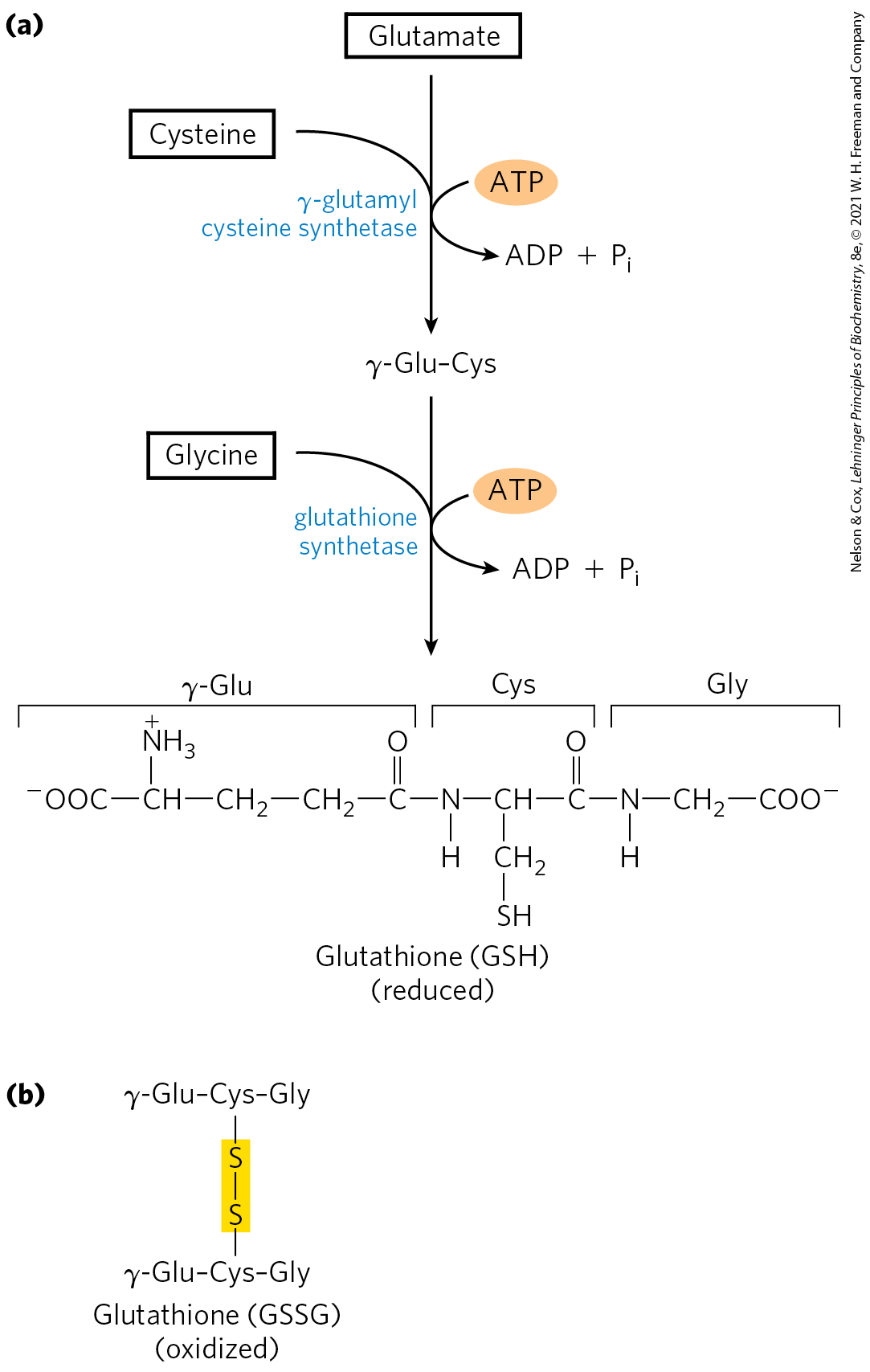
Glutathione (GSH), present in plants, animals, and some bacteria, often at high levels, can be thought of as a redox buffer. It is derived from glutamate, cysteine, and glycine (Fig. 22-29). The γ-carboxyl group of glutamate is activated by ATP to form an acyl phosphate intermediate, which is then attacked by the α-amino group of cysteine. A second condensation reaction follows, with the α-carboxyl group of cysteine activated to an acyl phosphate to permit reaction with glycine. The oxidized form of glutathione (glutathione disulfide, or GSSG), produced in the course of its redox activities, contains two glutathione molecules linked by a disulfide bond.
FIGURE 22-29 Glutathione metabolism. (a) Biosynthesis of glutathione. (b) Oxidized form of glutathione.
Glutathione helps maintain the sulfhydryl groups of proteins in the reduced state and the iron of heme in the ferrous state, and it serves as a reducing agent for glutaredoxin in deoxyribonucleotide synthesis (see Fig. 22-41). Its redox function is also used to remove toxic peroxides formed in the normal course of growth and metabolism under aerobic conditions:
This reaction is catalyzed by glutathione peroxidase, a remarkable enzyme in that it contains a covalently bound selenium (Se) atom in the form of selenocysteine (see Fig. 3-8a), which is essential for its activity.
d-Amino Acids Are Found Primarily in Bacteria
Although d-amino acids do not generally occur in proteins, they do serve some special functions in the structure of bacterial cell walls and peptide antibiotics. Bacterial peptidoglycans (see Fig. 6-32) contain both d-alanine and d-glutamate. d-Amino acids arise directly from the l isomers by the action of amino acid racemases, which have pyridoxal phosphate as cofactor (see Fig. 18-6). Amino acid racemization is uniquely important to bacterial metabolism, and enzymes such as alanine racemase are prime targets for pharmaceutical agents. One such agent, L-fluoroalanine, is being tested as an antibacterial drug. Another, cycloserine, is used to treat tuberculosis. Because these inhibitors also affect some PLP-requiring human enzymes, however, they have potentially undesirable side effects.
Aromatic Amino Acids Are Precursors of Many Plant Substances
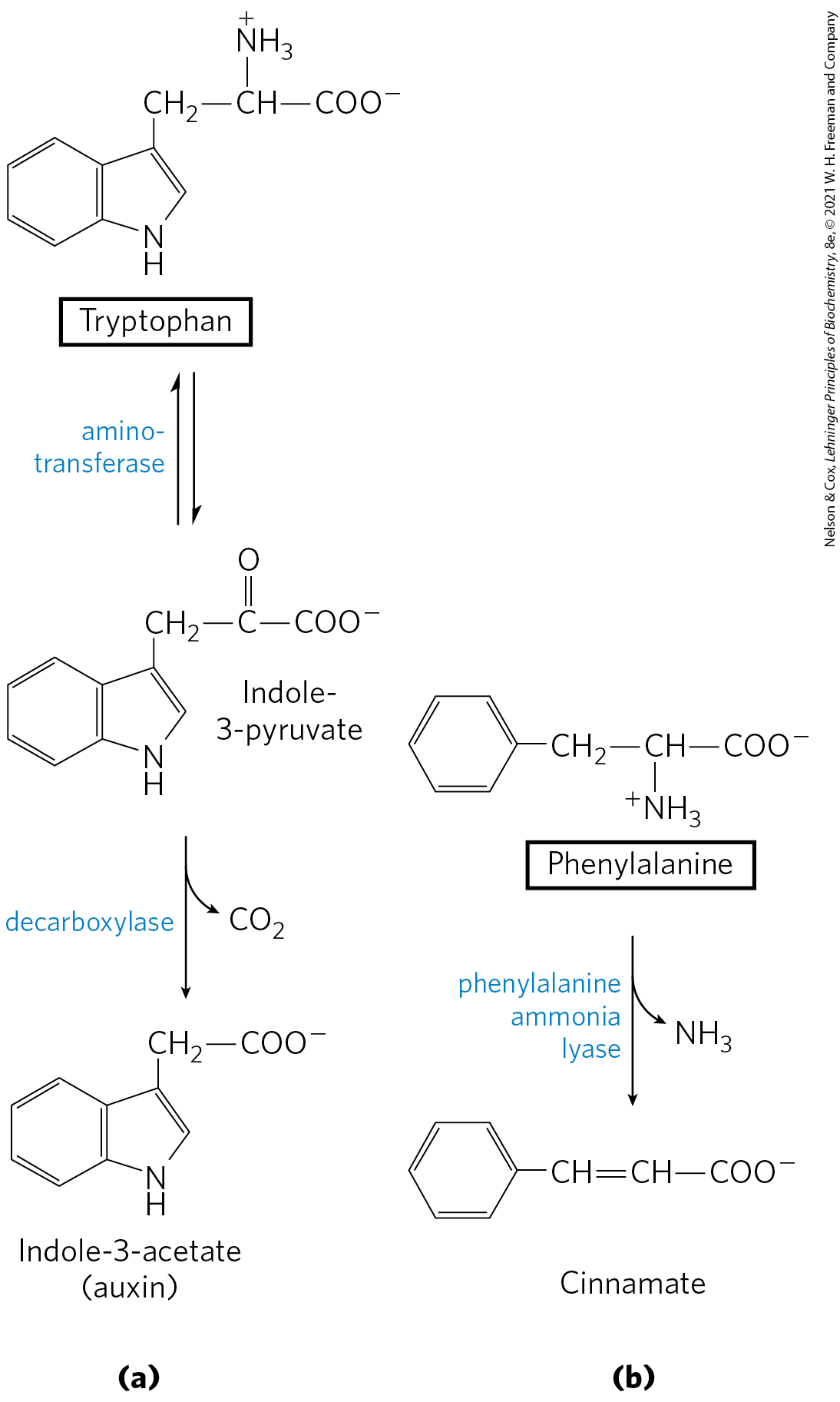
Phenylalanine, tyrosine, and tryptophan are converted to a variety of important compounds in plants. The rigid polymer lignin, derived from phenylalanine and tyrosine, is second only to cellulose in abundance in plant tissues. The structure of the lignin polymer is complex and not well understood. Tryptophan is also the precursor of the plant growth hormone indole-3-acetate, or auxin (Fig. 22-30a), which is important in the regulation of a wide range of biological processes in plants.
FIGURE 22-30 Biosynthesis of two plant substances from amino acids. (a) Indole-3-acetate (auxin) and (b) cinnamate (cinnamon flavor).
Phenylalanine and tyrosine also give rise to many commercially significant natural products, including the tannins that inhibit oxidation in wines; alkaloids such as morphine, which have potent physiological effects; and the flavoring of cinnamon oil (Fig. 22-30b), nutmeg, cloves, vanilla, cayenne pepper, and other products.
Biological Amines Are Products of Amino Acid Decarboxylation
Many important neurotransmitters are primary or secondary amines, derived from amino acids in simple pathways. In addition, some polyamines that form complexes with DNA are derived from the amino acid ornithine, a component of the urea cycle. A common denominator of many of these pathways is amino acid decarboxylation, another PLP-requiring reaction (see Fig. 18-6).
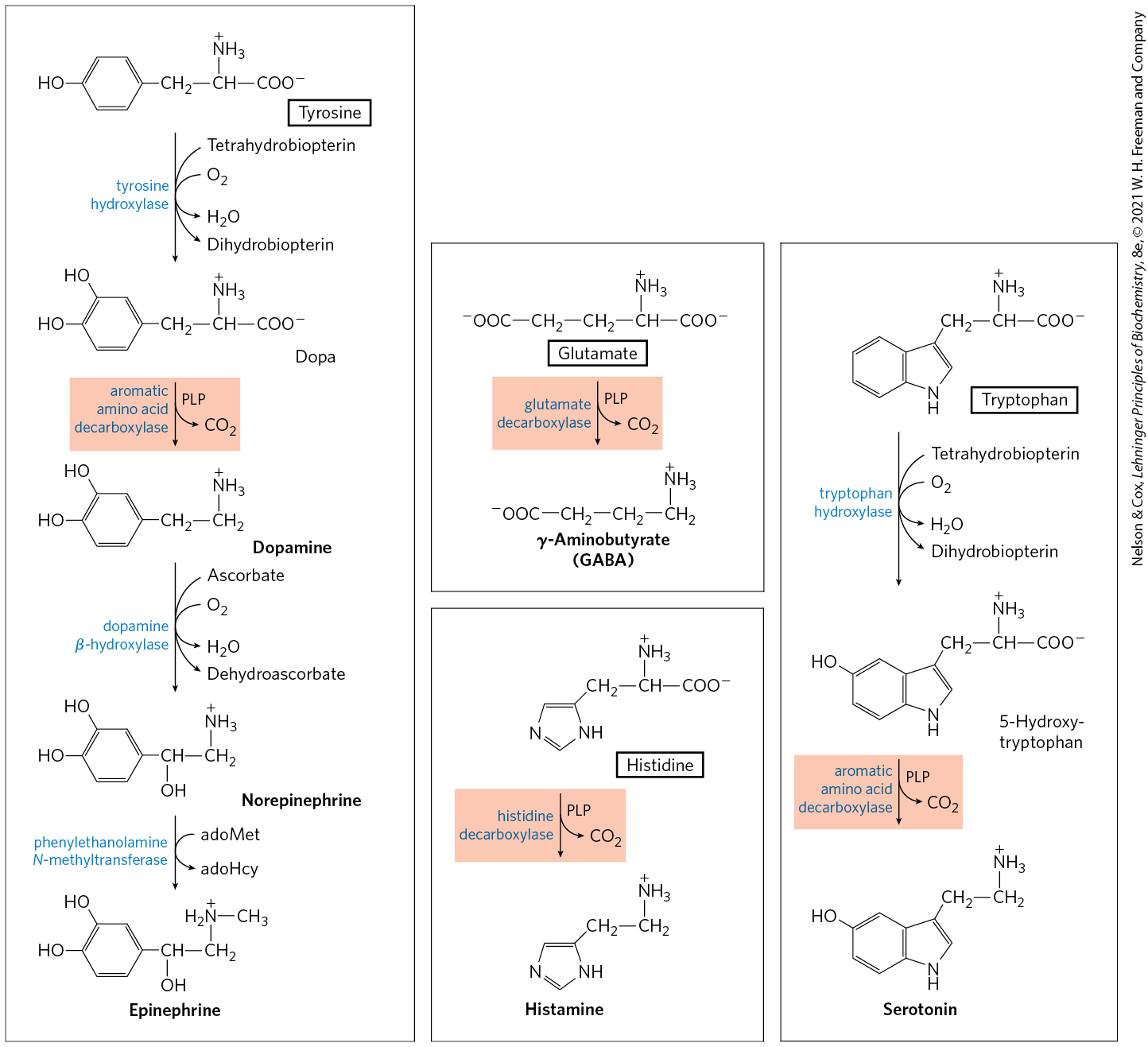
The synthesis of some neurotransmitters is illustrated in Figure 22-31. Tyrosine gives rise to a family of catecholamines that includes dopamine, norepinephrine, and epinephrine. Levels of catecholamines are correlated with, among other things, changes in blood pressure. The neurological disorder Parkinson disease is associated with an underproduction of dopamine, and it has traditionally been treated by administering l-dopa. Overproduction of dopamine in the brain may be linked to psychological disorders such as schizophrenia.
FIGURE 22-31 Biosynthesis of some neurotransmitters from amino acids. The key step is the same in each case: a PLP-dependent decarboxylation (shaded light red).
Glutamate decarboxylation gives rise to γ-aminobutyrate (GABA), an inhibitory neurotransmitter. Its underproduction is associated with epileptic seizures. GABA analogs are used in the treatment of epilepsy and hypertension. Levels of GABA can also be increased by administering inhibitors of the GABA-degrading enzyme GABA aminotransferase. Another important neurotransmitter, serotonin, is derived from tryptophan in a two-step pathway.
Histidine undergoes decarboxylation to histamine, a powerful vasodilator in animal tissues. Histamine is released in large amounts as part of the allergic response, and it also stimulates acid secretion in the stomach. A growing array of pharmaceutical agents are being designed to interfere with either the synthesis or the action of histamine. A prominent example is the histamine receptor antagonist cimetidine (Tagamet), a structural analog of histamine:
It promotes the healing of duodenal ulcers by inhibiting secretion of gastric acid.
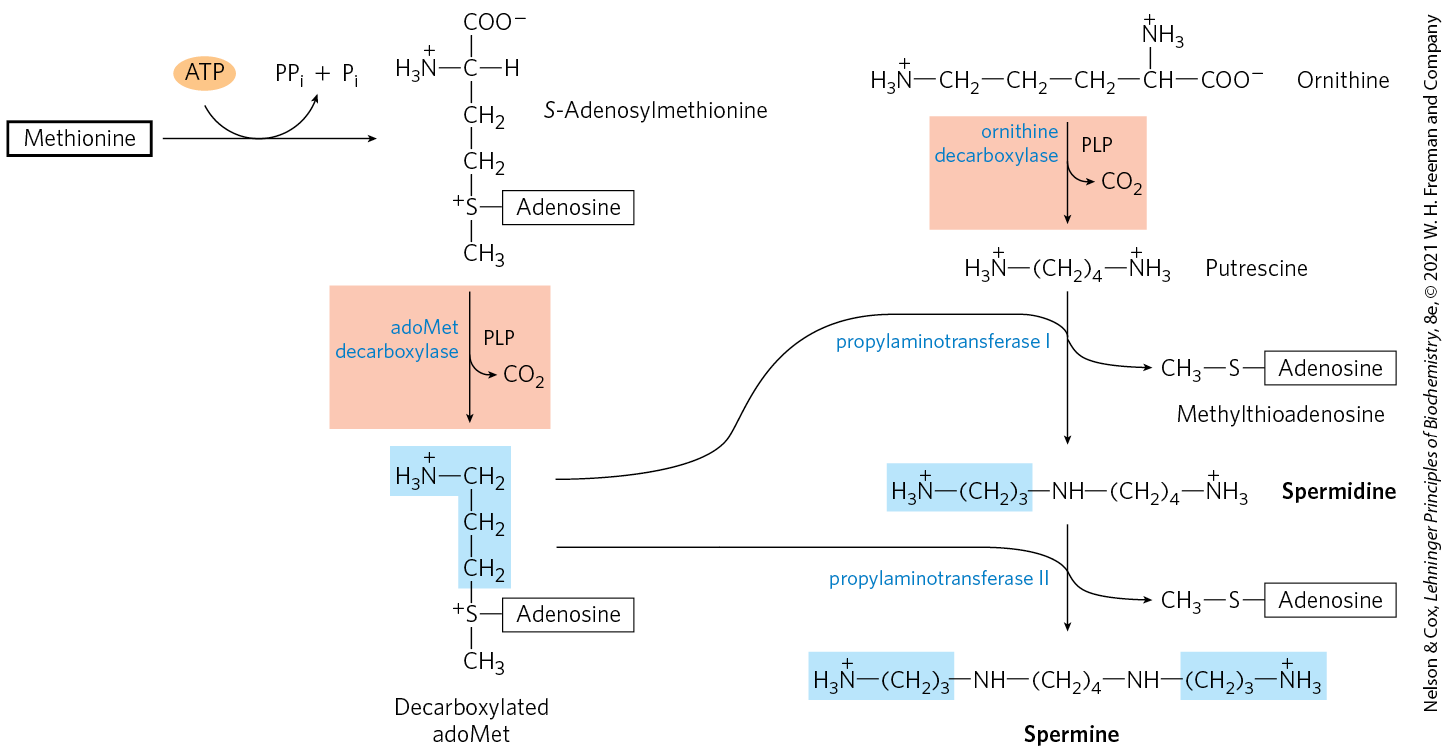
Polyamines such as spermine and spermidine, involved in DNA packaging, are derived from methionine and ornithine by the pathway shown in Figure 22-32. The first step is decarboxylation of ornithine, a precursor of arginine (Fig. 22-12). Ornithine decarboxylase, a PLP-requiring enzyme, is the target of several powerful inhibitors used as pharmaceutical agents (see Box 6-1).
FIGURE 22-32 Biosynthesis of spermidine and spermine. The PLP-dependent decarboxylation steps are shaded light red. In these reactions, S-adenosylmethionine (in its decarboxylated form) acts as a source of propylamino groups (shaded blue).
Arginine Is the Precursor for Biological Synthesis of Nitric Oxide
A surprise finding in the mid-1980s was the role of nitric oxide (NO) — previously known mainly as a component of smog — as an important biological messenger. This simple gaseous substance diffuses readily through membranes, although its high reactivity limits its range of diffusion to about a 1 mm radius from the site of synthesis. In humans NO plays a role in a range of physiological processes, including neurotransmission, blood clotting, and the control of blood pressure. Its mode of action is described in Box 12-2.
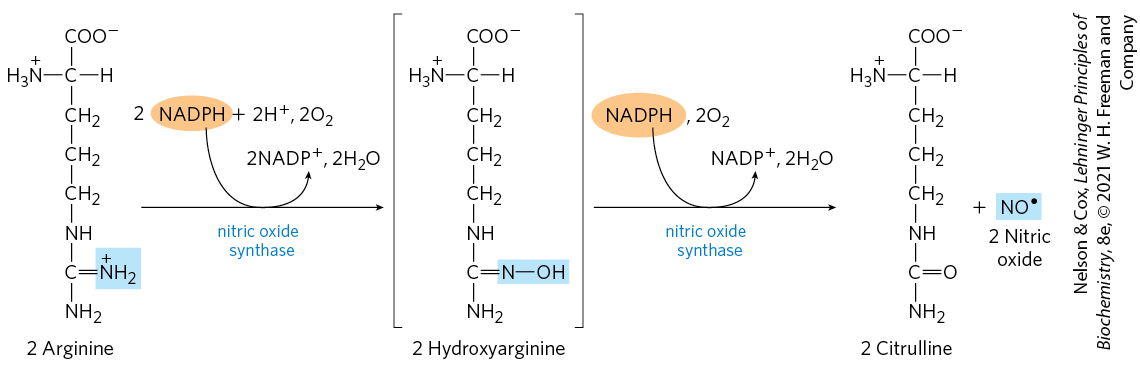
Nitric oxide is synthesized from arginine in an NADPH-dependent reaction catalyzed by nitric oxide synthase (Fig. 22-33), a dimeric enzyme structurally related to NADPH cytochrome P-450 reductase (see Box 21-1). The reaction is a five-electron oxidation. Each subunit of the enzyme contains one bound molecule of each of four different cofactors: FMN, FAD, tetrahydrobiopterin, and heme. NO is an unstable molecule and cannot be stored. Its synthesis is stimulated by interaction of nitric oxide synthase with -calmodulin (see Fig. 12-17).
FIGURE 22-33 Biosynthesis of nitric oxide. The nitrogen of the NO is derived from the guanidinium group of arginine.
SUMMARY 22.3 Molecules Derived from Amino Acids
Many important biomolecules are derived from amino acids. Glycine is a precursor of porphyrins.
Degradation of iron-porphyrin (heme) generates bilirubin, which is converted to bile pigments with several physiological functions.
Glycine and arginine give rise to creatine and phosphocreatine, an energy buffer. Glutathione, formed from three amino acids, is an important cellular reducing agent.
Bacteria synthesize d-amino acids from l-amino acids in racemization reactions requiring pyridoxal phosphate. d-Amino acids are commonly found in certain bacterial walls and certain antibiotics.
Plants make many substances from aromatic amino acids.
The PLP-dependent decarboxylation of some amino acids yields important biological amines, including neurotransmitters and polyamines.
Arginine is the precursor of nitric oxide, a biological messenger.
 reduction by NADPH converts the glutamate to glutamate 1-semialdehyde, which is cleaved from the tRNA. An aminotransferase converts the glutamate 1-semialdehyde to δ-aminolevulinate.
reduction by NADPH converts the glutamate to glutamate 1-semialdehyde, which is cleaved from the tRNA. An aminotransferase converts the glutamate 1-semialdehyde to δ-aminolevulinate.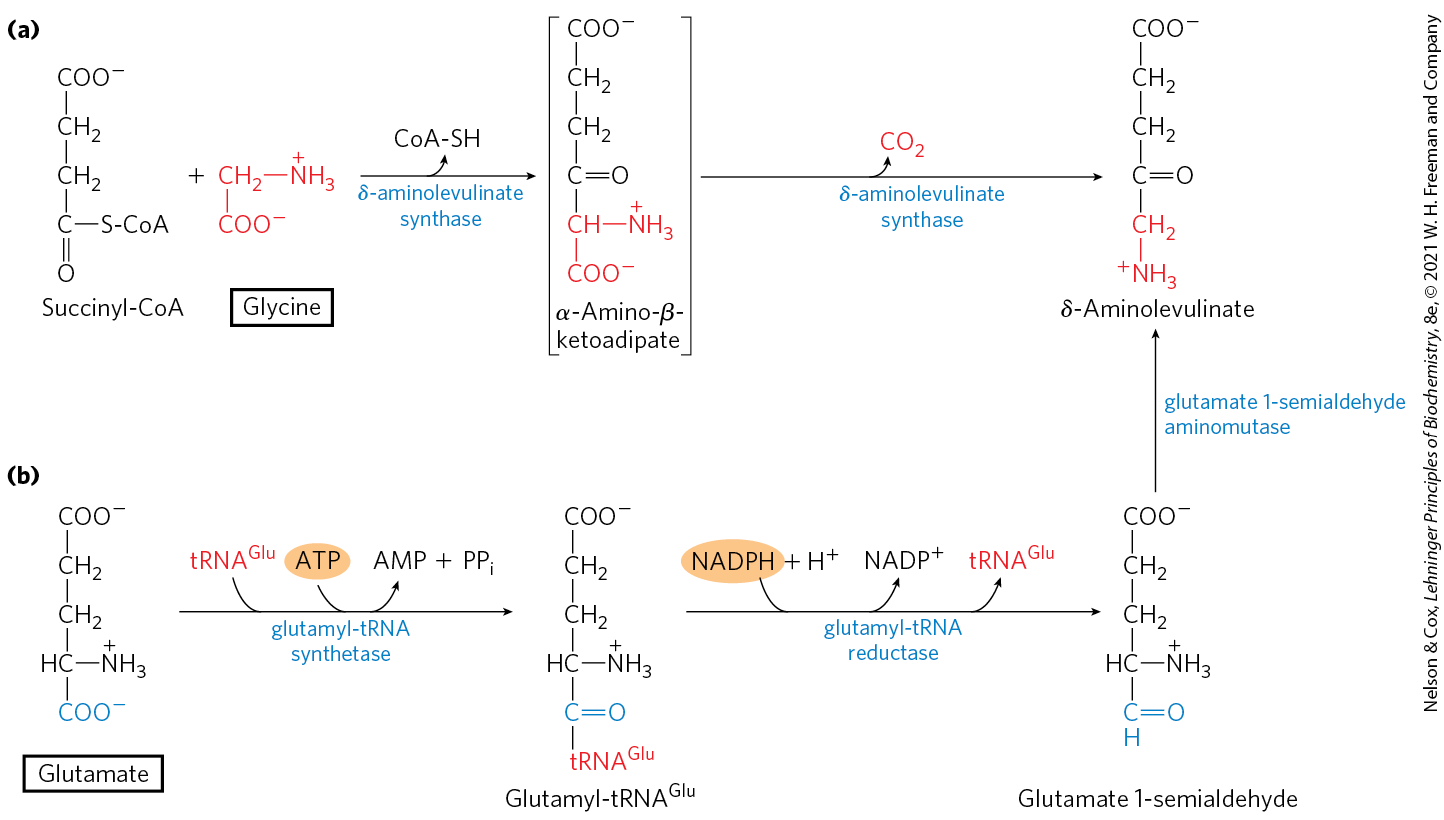
 The iron-porphyrin (heme) group of hemoglobin, released from dying erythrocytes in the spleen, is degraded to yield free and, ultimately, bilirubin. The pathway also contributes the pigment present in mixtures of the bile salts derived from cholesterol.
The iron-porphyrin (heme) group of hemoglobin, released from dying erythrocytes in the spleen, is degraded to yield free and, ultimately, bilirubin. The pathway also contributes the pigment present in mixtures of the bile salts derived from cholesterol.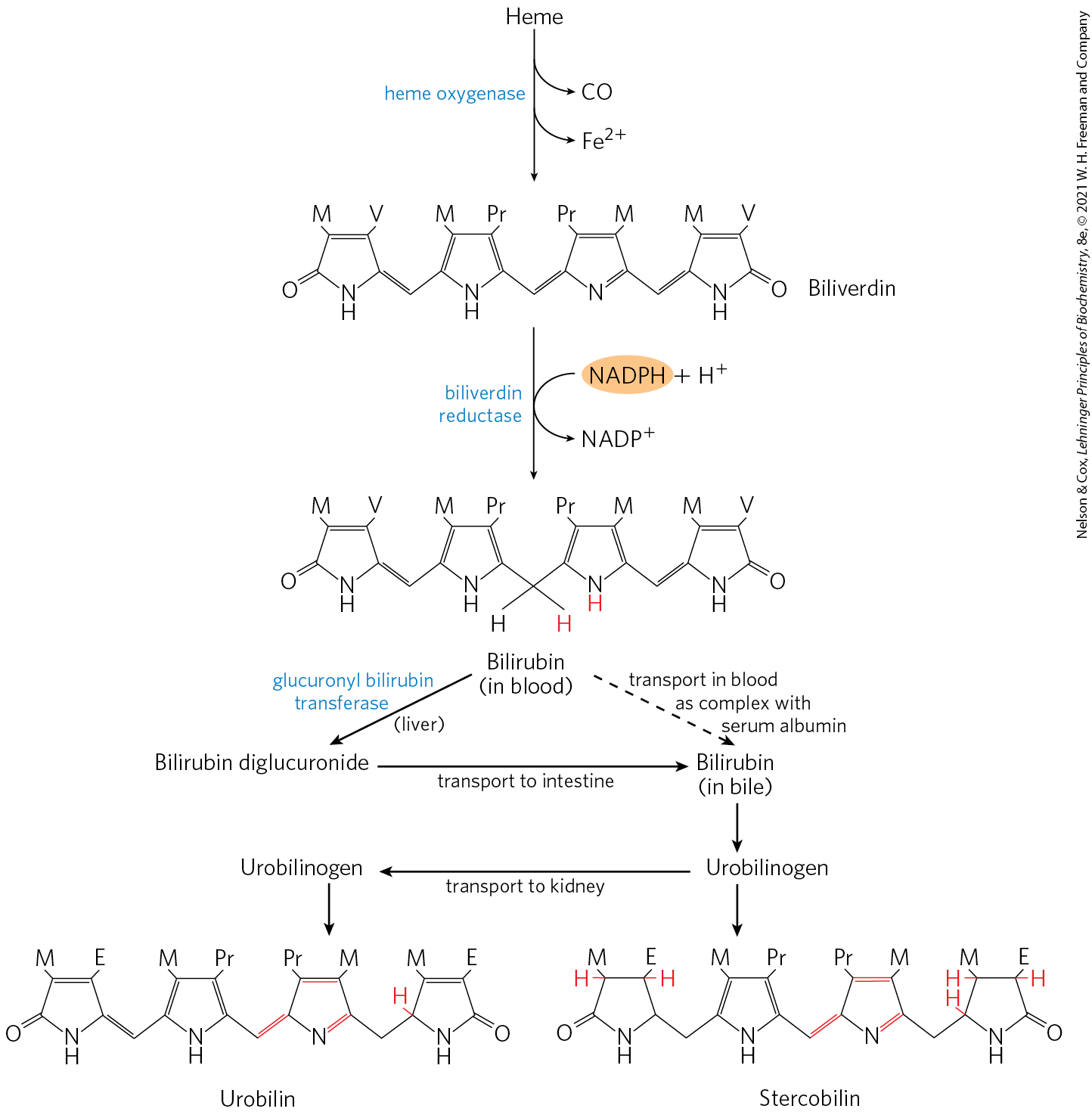
 Given these varied roles of heme degradation products, the degradative pathway is subject to regulation, mainly at the first step. Humans have at least three isozymes of heme oxygenase (HO). HO-1 is highly regulated; the expression of its gene is induced by a wide range of stress conditions, including shear stress, uncontrolled angiogenesis (development of blood vessels), hypoxia, hyperoxia, heat shock, exposure to ultraviolet light, hydrogen peroxide, and many other metabolic insults. HO-2 is found mainly in the brain and the testes, where it is continuously expressed. The third isozyme, HO-3, is not catalytically active, but may play a role in oxygen sensing.
Given these varied roles of heme degradation products, the degradative pathway is subject to regulation, mainly at the first step. Humans have at least three isozymes of heme oxygenase (HO). HO-1 is highly regulated; the expression of its gene is induced by a wide range of stress conditions, including shear stress, uncontrolled angiogenesis (development of blood vessels), hypoxia, hyperoxia, heat shock, exposure to ultraviolet light, hydrogen peroxide, and many other metabolic insults. HO-2 is found mainly in the brain and the testes, where it is continuously expressed. The third isozyme, HO-3, is not catalytically active, but may play a role in oxygen sensing. 
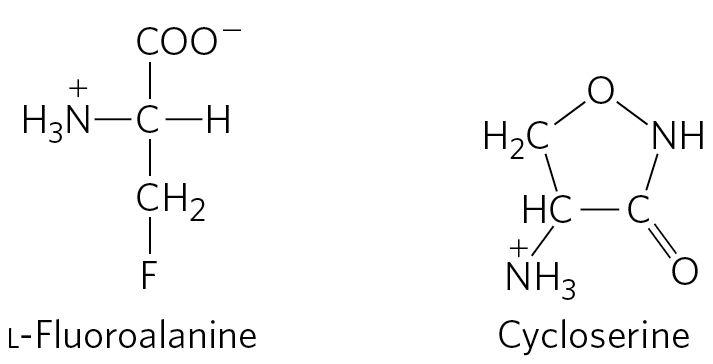
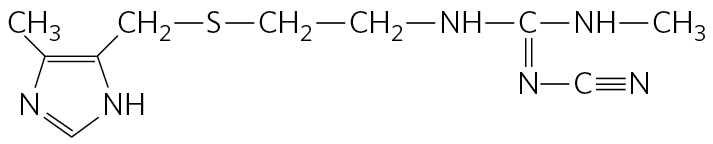
 Many important biomolecules are derived from amino acids. Glycine is a precursor of porphyrins.
Many important biomolecules are derived from amino acids. Glycine is a precursor of porphyrins.