Chapter Review
KEY TERMS
Terms in bold are defined in the glossary.
- autotroph
- heterotroph
- metabolite
- intermediary metabolism
- catabolism
- anabolism
- energy transduction
- free energy, G
- exergonic
- endergonic
- enthalpy, H
- exothermic
- endothermic
- entropy, S
- standard transformed constants
- mass-action ratio, Q
- homolytic cleavage
- radical
- heterolytic cleavage
- nucleophile
- electrophile
- carbanion
- carbocation
- aldol condensation
- Claisen condensation
- kinases
- phosphorylation potential
- thioester
- adenylylation
- inorganic pyrophosphatase
- nucleoside diphosphate kinase
- adenylate kinase
- creatine kinase
- phosphagens
- electromotive force (emf)
- conjugate redox pair
- dehydrogenases
- reducing equivalent
- standard reduction potential
- pyridine nucleotide
- oxidoreductase
- flavoprotein
- flavin nucleotides
- glucose 6-phosphate
- homeostasis
- transcription factor
- response element
- turnover
- transcriptome
- proteome
- metabolome
- metabolic regulation
- metabolic control
- adenylate kinase
- AMP-activated protein kinase (AMPK)
Problems
1. Entropy Changes during Egg Development Consider a system consisting of an egg in an incubator. The white and yolk of the egg contain proteins, carbohydrates, and lipids. If fertilized, the egg transforms from a single cell to a complex organism. Discuss this irreversible process in terms of the entropy changes in the system and surroundings. Be sure that you first clearly define the system and surroundings.
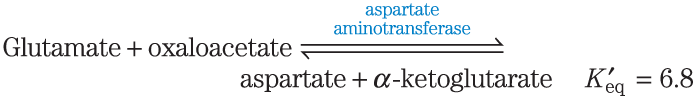
2. Calculation of from an Equilibrium Constant Calculate the standard free-energy change for each of the three metabolically important enzyme-catalyzed reactions, using the equilibrium constants given for the reactions at and pH 7.0.
3. Calculation of the Equilibrium Constant from Calculate the equilibrium constant for each of the three reactions at pH 7.0 and , using the values in Table 13-4.
4. Experimental Determination of and Incubating a 0.1 m solution of glucose 1-phosphate at with a catalytic amount of phosphoglucomutase transforms some of the glucose 1-phosphate to glucose 6-phosphate. At equilibrium, the concentrations of the reaction components are
Calculate and for this reaction.
5. Experimental Determination of for ATP Hydrolysis A direct measurement of the standard free-energy change associated with the hydrolysis of ATP is technically demanding because the minute amount of ATP remaining at equilibrium is difficult to measure accurately. The value of can be calculated indirectly, however, from the equilibrium constants of two other enzymatic reactions having less favorable equilibrium constants:
Using this information for equilibrium constants determined at , calculate the standard free energy of hydrolysis of ATP.
6. Difference between and ΔG Consider the interconversion shown, which occurs in glycolysis (Chapter 14):
- What is for the reaction ( measured at )?
- If the concentration of fructose 6-phosphate is adjusted to 1.5 m and that of glucose 6-phosphate is adjusted to 0.50 m, what is ΔG?
- Why are and ΔG different?
7. Free Energy of Hydrolysis of CTP Compare the structure of the nucleoside triphosphate CTP with the structure of ATP.
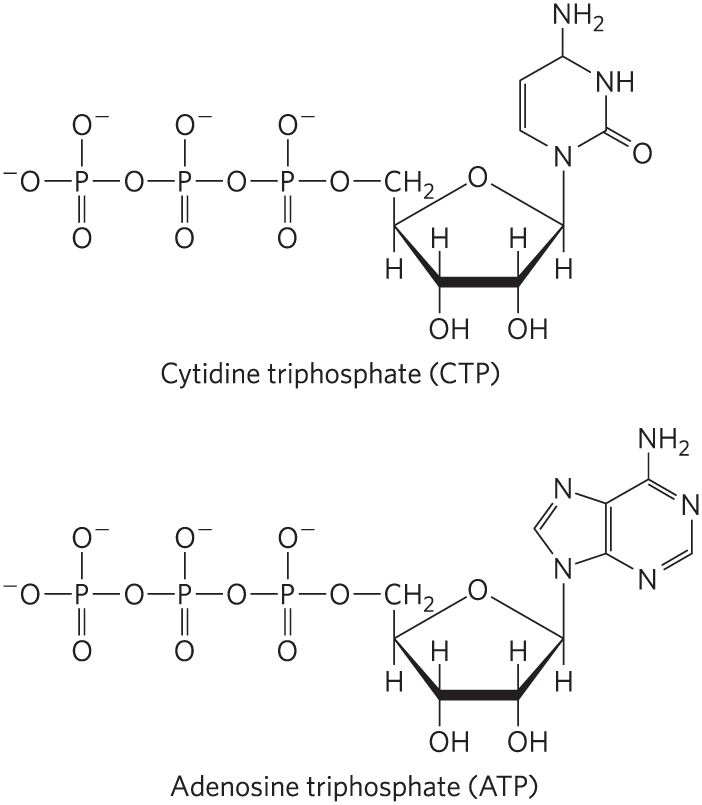
Now predict the and for the reaction:
8. Dependence of ΔG on pH The free energy released by the hydrolysis of ATP under standard conditions is . If ATP is hydrolyzed under standard conditions except at pH 5.0, is more or less free energy released? Explain.
9. The for Coupled Reactions Glucose 1-phosphate is converted into fructose 6-phosphate in two successive reactions:
Using the values in Table 13-4, calculate the equilibrium constant, , for the sum of the two reactions:
10. Effect of [ATP]/[ADP] Ratio on Free Energy of Hydrolysis of ATP Using Equation 13-4, plot ΔG against ln Q (mass-action ratio) at for the concentrations of ATP, ADP, and in the table shown. for the reaction is . Use the resulting plot to explain why metabolism is regulated to keep the ratio [ATP]/[ADP] high.
Concentration (mm) ATP 5 3 1 0.2 5 ADP 0.2 2.2 4.2 5.0 25 10 12.1 14.1 14.9 10 11. Strategy for Overcoming an Unfavorable Reaction: ATP-Dependent Chemical Coupling The phosphorylation of glucose to glucose 6-phosphate is the initial step in the catabolism of glucose. The direct phosphorylation of glucose by is described by the equation
- Calculate the equilibrium constant for this reaction at . In the rat hepatocyte, the physiological concentrations of glucose and are maintained at approximately 4.8 mm. What is the equilibrium concentration of glucose 6-phosphate obtained by the direct phosphorylation of glucose by ? Does this reaction represent a reasonable metabolic step for the catabolism of glucose? Explain.
- In principle at least, one way to increase the concentration of glucose 6-phosphate is to drive the equilibrium reaction to the right by increasing the intracellular concentrations of glucose and . Assuming a fixed concentration of at 4.8 mm, how high would the intracellular concentration of glucose have to be to give an equilibrium concentration of glucose 6-phosphate of 250 μm (the normal physiological concentration)? Would this route be physiologically reasonable, given that the maximum solubility of glucose is less than 1 m?
- The phosphorylation of glucose in the cell is coupled to the hydrolysis of ATP; that is, part of the free energy of ATP hydrolysis is used to phosphorylate glucose:
Calculate at for the overall reaction. For the ATP-dependent phosphorylation of glucose, what concentration of glucose is needed to achieve a 250 μm intracellular concentration of glucose 6-phosphate when the concentrations of ATP and ADP are 3.38 mm and 1.32 mm, respectively? Does this coupling process provide a feasible route, at least in principle, for the phosphorylation of glucose in the cell? Explain.
- Although coupling ATP hydrolysis to glucose phosphorylation makes thermodynamic sense, we have not yet specified how this coupling is to take place. Given that coupling requires a common intermediate, one conceivable route is to use ATP hydrolysis to raise the intracellular concentration of and thus drive the unfavorable phosphorylation of glucose by . Is this a reasonable route? (Think about the solubility product, , of metabolic intermediates.)
- The ATP-coupled phosphorylation of glucose is catalyzed in hepatocytes by the enzyme glucokinase. This enzyme binds ATP and glucose to form a glucose-ATP-enzyme complex, and the phosphoryl group is transferred directly from ATP to glucose. Explain the advantages of this route.
12. Calculations of for ATP-Coupled Reactions From data in Table 13-6, calculate the value for each reaction:
13. Coupling ATP Cleavage to an Unfavorable Reaction To explore the consequences of coupling ATP hydrolysis under physiological conditions to a thermodynamically unfavorable biochemical reaction, consider the hypothetical transformation , for which .
- What is the ratio [Y]/[X] at equilibrium?
- Suppose X and Y participate in a sequence of reactions during which ATP is hydrolyzed to ADP and . The overall reaction is
Calculate [Y]/[X] for this reaction at equilibrium. Assume that the temperature is and the equilibrium concentrations of ATP, ADP, and are 1 m.
- We know that [ATP], [ADP], and are not 1 m under physiological conditions. Calculate [Y]/[X] for the ATP-coupled reaction when the values of [ATP], [ADP], and are those found in rat myocytes (Table 13-5).
14. Calculations of ΔG at Physiological Concentrations Calculate the actual, physiological ΔG for the reaction
at , as it occurs in the cytosol of neurons, with phosphocreatine at 4.7 mm, creatine at 1.0 mm, ADP at 0.73 mm, and ATP at 2.6 mm.
15. Free Energy Required for ATP Synthesis under Physiological Conditions In the cytosol of rat hepatocytes, the temperature is and the mass-action ratio, Q, is
Calculate the free energy required to synthesize ATP in a rat hepatocyte.
16. Chemical Logic In the glycolytic pathway, a six-carbon sugar (fructose 1,6-bisphosphate) is cleaved to form two three-carbon sugars, which undergo further metabolism. In this pathway, an isomerization of glucose 6-phosphate to fructose 6-phosphate (as shown in the diagram) occurs two steps before the cleavage reaction. The intervening step is phosphorylation of fructose 6-phosphate to fructose 1,6-bisphosphate (p. 516).

What does the isomerization step accomplish from a chemical perspective? (Hint: Consider what might happen if the bond cleavage were to proceed without the preceding isomerization.)
17. Enzymatic Reaction Mechanisms I Lactate dehydrogenase is one of the many enzymes that require NADH as coenzyme. It catalyzes the conversion of pyruvate to lactate:
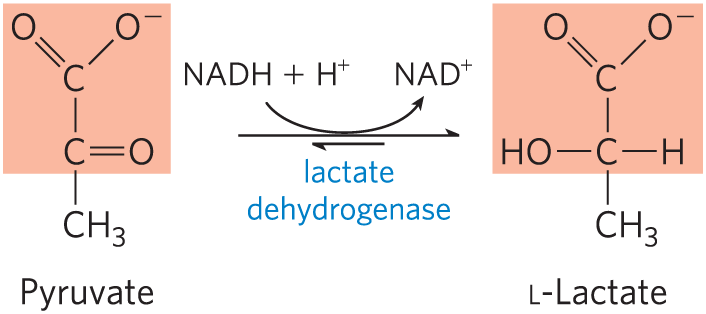
Draw the mechanism of this reaction (show electron-pushing arrows). (Hint: This is a common reaction throughout metabolism; the mechanism is similar to that catalyzed by other dehydrogenases that use NADH, such as alcohol dehydrogenase.)
18. Enzymatic Reaction Mechanisms II Biochemical reactions often look more complex than they really are. In the pentose phosphate pathway (Chapter 14), sedoheptulose 7-phosphate and glyceraldehyde 3-phosphate react to form erythrose 4-phosphate and fructose 6-phosphate in a reaction catalyzed by transaldolase.
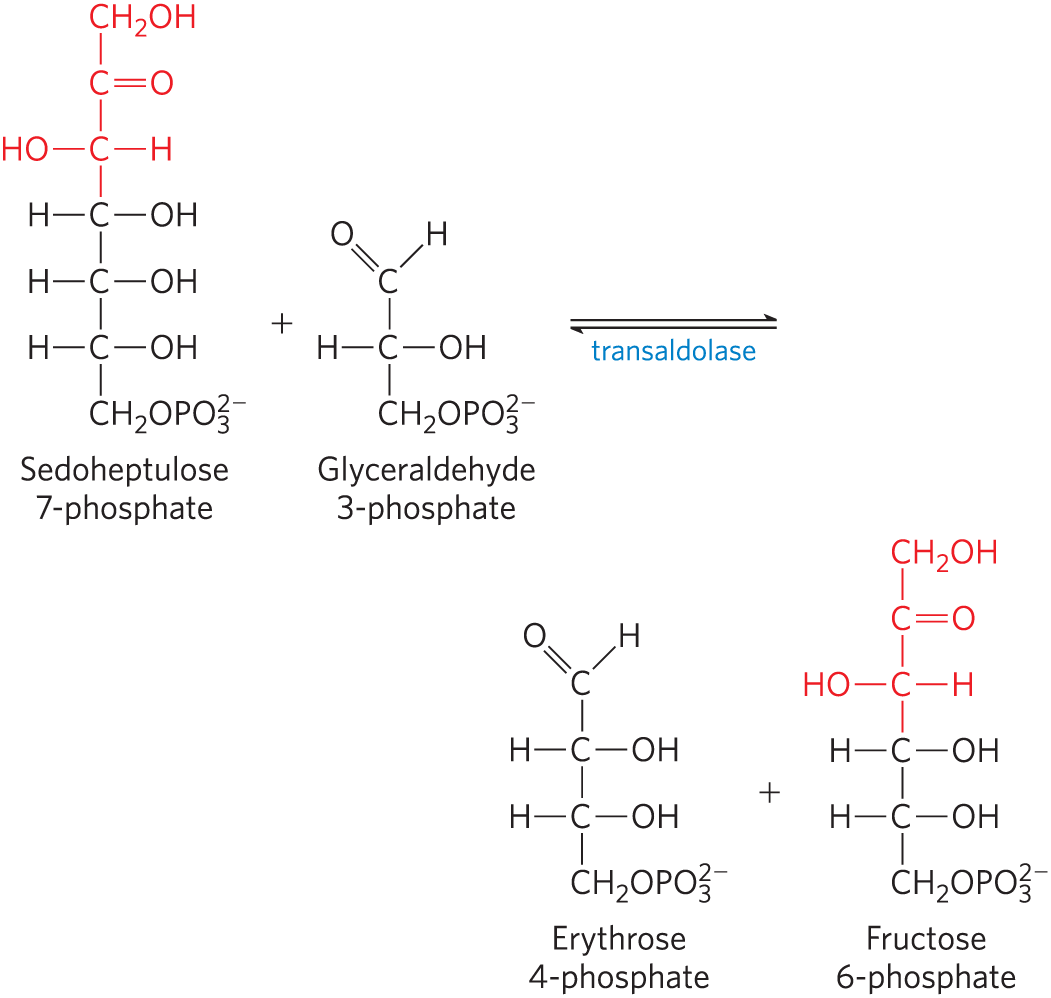
Draw a mechanism for this reaction (show electron-pushing arrows). (Hint: Take another look at aldol condensations, then consider the name of this enzyme.)
19. Recognizing Reaction Types For each pair of biomolecules, identify the type of reaction (oxidation-reduction, hydrolysis, isomerization, group transfer, or internal rearrangement) required to convert the first molecule to the second. In each case, indicate the general type of enzyme and cofactor(s) or reactants that would be required, and any other products that would result.
-
20. Effect of Structure on Group Transfer Potential Some invertebrates contain phosphoarginine. Is the standard free energy of hydrolysis of this molecule more similar to that of glucose 6-phosphate or of ATP? Explain your answer.
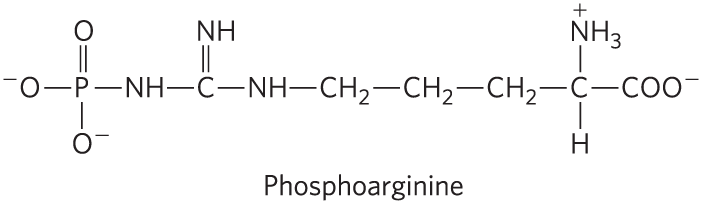
21. Polyphosphate as a Possible Energy Source The standard free energy of hydrolysis of inorganic polyphosphate (polyP) is about for each released. We calculated in Worked Example 13-2 that, in a cell, it takes about 50 kJ/mol of energy to synthesize ATP from ADP and .
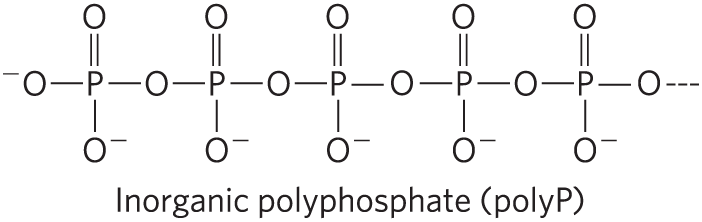
Is it feasible for a cell to use polyphosphate to synthesize ATP from ADP? Explain your answer.
22. Daily ATP Utilization by Human Adults
- The synthesis of ATP from ADP and requires a total of 30.5 kJ/mol of free energy when the reactants and products are at 1 m concentrations and the temperature is (standard state). However, the actual physiological concentrations of ATP, ADP, and are not 1 m, and the physiological temperature is 37 °C. Thus, the free energy required to synthesize ATP under physiological conditions is different from . Calculate the free energy required to synthesize ATP in the human hepatocyte when the physiological concentrations of ATP, ADP, and are 3.5, 1.50, and 5.0 mm, respectively.
- A 68 kg (150 lb) adult requires a caloric intake of 2,000 kcal (8,360 kJ) of food per day (24 hours). The body metabolizes the food and uses the free energy to synthesize ATP, which then provides energy for the body’s daily chemical and mechanical work. Assuming that the efficiency of converting food energy into ATP is 50%, calculate the weight of ATP used by a human adult in 24 hours. What percentage of the body weight does this represent?
- Although adults synthesize large amounts of ATP daily, their body weight, structure, and composition do not change significantly during this period. Explain this apparent contradiction.
23. Rates of Turnover of γ and β Phosphates of ATP After adding a small amount of ATP labeled with radioactive phosphorus in the terminal position, ATP, to a yeast extract, a researcher finds about half of the activity in within a few minutes, but the concentration of ATP remains unchanged. Explain. She then carries out the same experiment using ATP labeled with in the central position, ATP, but the does not appear in within such a short time. Why?
24. Cleavage of ATP to AMP and during Metabolism Synthesis of the activated form of acetate (acetyl-CoA) is carried out in an ATP-dependent process:
- The for hydrolysis of acetyl-CoA to acetate and CoA is . The for hydrolysis of ATP to AMP and is . Calculate for the ATP-dependent synthesis of acetyl-CoA.
- Almost all cells contain the enzyme inorganic pyrophosphatase, which catalyzes the hydrolysis of to . What effect does the presence of this enzyme have on the synthesis of acetyl-CoA? Explain.
25. Activation of a Fatty Acid by Reaction with Coenzyme A In the reaction sequence for fatty acid breakdown, coenzyme A (CoA), with its thiol group, joins to the fatty acid as a thiol ester, as ATP is converted into AMP and :
The oxidation of fatty acids as fuels requires two steps. The first step transfers an activating group from ATP to the carboxyl group of the fatty acid. In the second step, displaces the activating group to form fatty acyl-. Given the known products of the reaction, what is the activating group?
26. Energy for Pumping The parietal cells of the stomach lining contain membrane “pumps” that transport hydrogen ions from the cytosol (pH 7.0) into the stomach, contributing to the acidity of gastric juice (pH 1.0). Calculate the free energy required to transport 1 mol of hydrogen ions through these pumps. (Hint: See Chapter 11.) Assume a temperature of 37 °C.
27. Most-Reduced Carbon Compounds Arrange the four structures in order from most reduced to most oxidized.
28. Standard Reduction Potentials The standard reduction potential, , of any redox pair is defined for the half-cell reaction
The values for the and pyruvate/lactate conjugate redox pairs are and respectively.
- Which redox pair has the greater tendency to lose electrons? Explain.
- Which pair is the stronger oxidizing agent? Explain.
- Beginning with 1 m concentrations of each reactant and product at pH 7 and 25 °C, in which direction will the following reaction proceed?
- What is the standard free-energy change for the conversion of pyruvate to lactate?
- What is the equilibrium constant () for this reaction?
29. Simple Biobattery Suppose you set up a simple battery using half-reactions as pictured in Figure 13-23. One electrode contains pyruvate and lactate at 1 mm, and the other electrode contains fumarate and succinate at 1 mm (see Table 13-7).
- In which direction will electrons initially flow?
- Calculate the standard reduction potential and standard free-energy change for your biological battery.
- When a flashlight battery “runs out,” net electron movement has essentially ended. What is the equivalent situation for your biobattery?
30. Energy Span of the Respiratory Chain Electron transfer in the mitochondrial respiratory chain may be represented by the net reaction equation
- Calculate for the net reaction of mitochondrial electron transfer. Use values in Table 13-7.
- Calculate for this reaction.
- How many ATP molecules can theoretically be generated by this reaction if the free energy of ATP synthesis under cellular conditions is 52 kJ/mol?
31. Dependence of Electromotive Force on Concentrations Suppose that you place an electrode into solutions of various concentrations of and NADH at pH 7.0 and 25 °C. Calculate the electromotive force (in volts) registered by the electrode when immersed in each solution, with reference to a half-cell of 0.00 V.
- 1.0 mm and 10 mm NADH
- 1.0 mm and 1.0 mm NADH
- 10 mm and 1.0 mm NADH
32. Electron Affinity of Compounds List the four compounds or reactions in order of increasing tendency to accept electrons:
- (yielding isocitrate)
- oxaloacetate
33. Direction of Oxidation-Reduction Reactions Which of the reactions listed would you expect to proceed in the direction shown, under standard conditions, in the presence of the appropriate enzymes?
34. Measurement of Intracellular Metabolite Concentrations Measuring the concentrations of metabolic intermediates in a living cell presents great experimental difficulties — usually, a cell must be destroyed before metabolite concentrations can be measured. Yet enzymes catalyze metabolic interconversions very rapidly, so a common problem associated with these types of measurements is that the findings reflect not the physiological concentrations of metabolites but the equilibrium concentrations. To prevent changes in metabolite concentrations during sample preparation, cells were quick-frozen in liquid nitrogen, then extracted under conditions that prevented enzymatic activity.
The table gives the intracellular concentrations of the substrates and products of the phosphofructokinase-1 reaction in isolated rat heart tissue.
Metabolite Concentration (μM)a Fructose 6-phosphate 87.0 Fructose 1,6-bisphosphate 22.0 ATP 11,400 ADP 1,320 Data from J. R. Williamson, J. Biol. Chem. 240:2308, 1965.
aCalculated as μmol/mL of intracellular water.
- Calculate Q, [fructose 1,6-bisphosphate][ADP]/[fructose 6-phosphate][ATP], for the PFK-1 reaction under physiological conditions.
- Given a for the PFK-1 reaction of , calculate the equilibrium constant for this reaction.
- Compare the values of Q and . Is the physiological reaction near or far from equilibrium? Explain. What does this experiment suggest about the role of PFK-1 as a regulatory enzyme?
35. Are All Metabolic Reactions at Equilibrium?
- Phosphoenolpyruvate (PEP) is one of the two phosphoryl group donors in the synthesis of ATP during glycolysis. In human erythrocytes, the steady-state concentration of ATP is 2.24 mm, that of ADP is 0.25 mm, and that of pyruvate is 0.051 mm. Calculate the concentration of PEP at 25 °C, assuming that the pyruvate kinase reaction (see Fig. 13-13) is at equilibrium in the cell.
- The physiological concentration of PEP in human erythrocytes is 0.023 mm. Compare this with the value obtained in (a). Explain the significance of this difference.
36. Michaelis Constant Compared with Substrate Concentration Malate synthase in E. coli catalyzes the reaction
The experimentally measured for acetyl-CoA is . In a growing culture of E. coli, the measured concentration of acetyl-CoA is . Is malate synthase operating at its under these conditions?
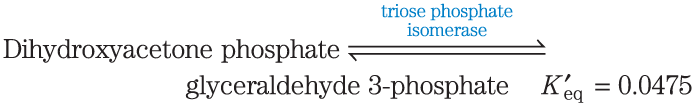
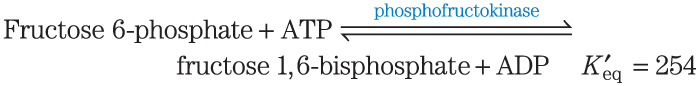


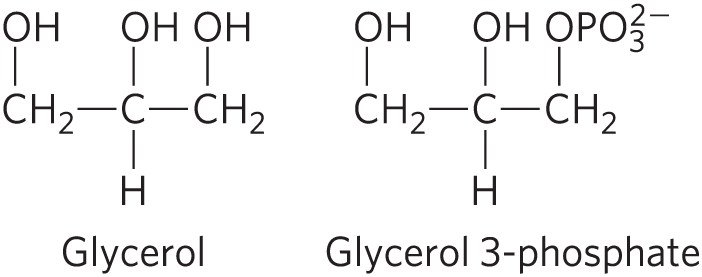
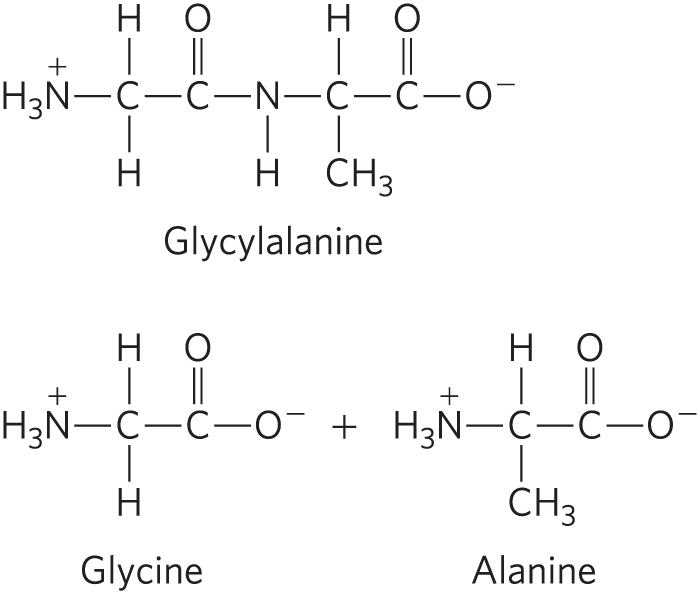
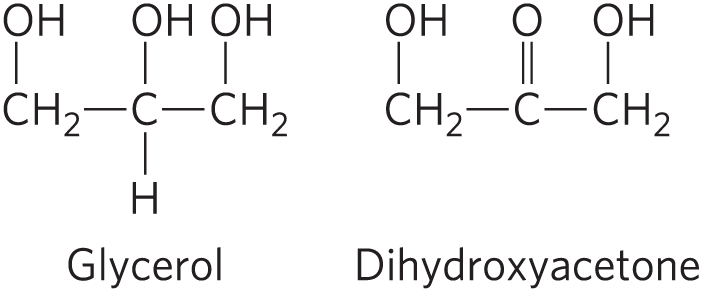
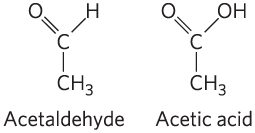
DATA ANALYSIS PROBLEM
37. Thermodynamics Can Be Tricky Thermodynamics is a challenging area of study and one with many opportunities for confusion. An interesting example is found in an article by Robinson, Hampson, Munro, and Vaney, published in Science in 1993. Robinson and colleagues studied the movement of small molecules between neighboring cells of the nervous system through cell-to-cell channels (gap junctions). They found that the dyes Lucifer yellow (a small, negatively charged molecule) and biocytin (a small zwitterionic molecule) moved in only one direction between two particular types of glia (nonneuronal cells of the nervous system). Dye injected into astrocytes would rapidly pass into adjacent astrocytes, oligodendrocytes, or Müller cells, but dye injected into oligodendrocytes or Müller cells passed slowly if at all into astrocytes. All of these cell types are connected by gap junctions.
Although it was not a central point of their article, the authors presented a molecular model for how this unidirectional transport might occur, as shown in their Figure 3:
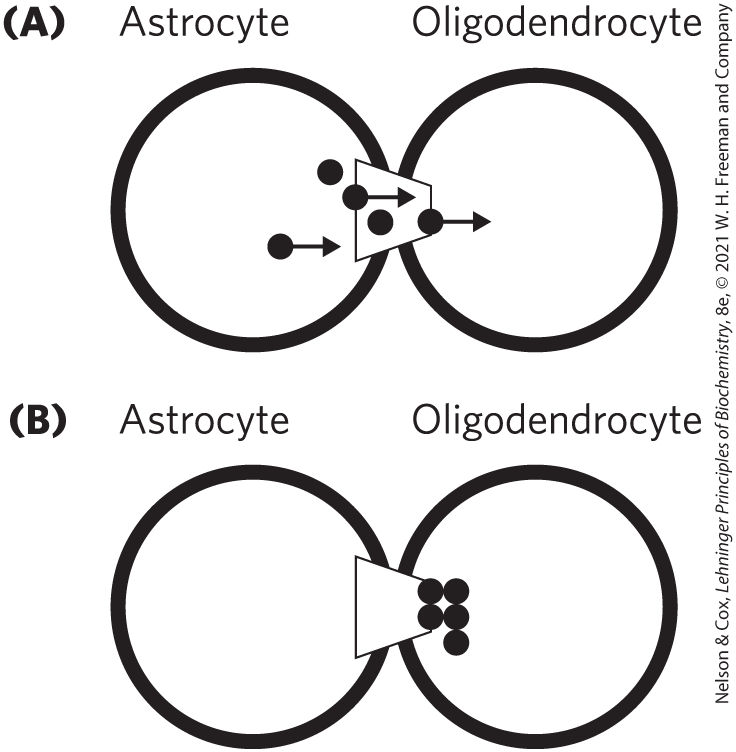
The figure legend reads: “Model of the unidirectional diffusion of dye between coupled oligodendrocytes and astrocytes, based on differences in connection pore diameter. Like a fish in a fish trap, dye molecules (black circles) can pass from an astrocyte to an oligodendrocyte (A) but not back in the other direction (B).”
Although this article clearly passed review at a well-respected journal, several letters to the editor (1994) followed, showing that Robinson and coauthors’ model violated the second law of thermodynamics.
- Explain how the model violates the second law. Hint: Consider what would happen to the entropy of the system if one started with equal concentrations of dye in the astrocyte and oligodendrocyte connected by the “fish trap” type of gap junctions.
- Explain why this model cannot work for small molecules, although it may allow one to catch fish.
- Explain why a fish trap does work for fish.
- Provide two plausible mechanisms for the unidirectional transport of dye molecules between the cells that do not violate the second law of thermodynamics.
References
- Letters to the editor. 1994. Science 265:1017–1019.
- Robinson, S.R., E.C.G.M. Hampson, M.N. Munro, and D.I. Vaney. 1993. Unidirectional coupling of gap junctions between neuroglia. Science 262:1072–1074.