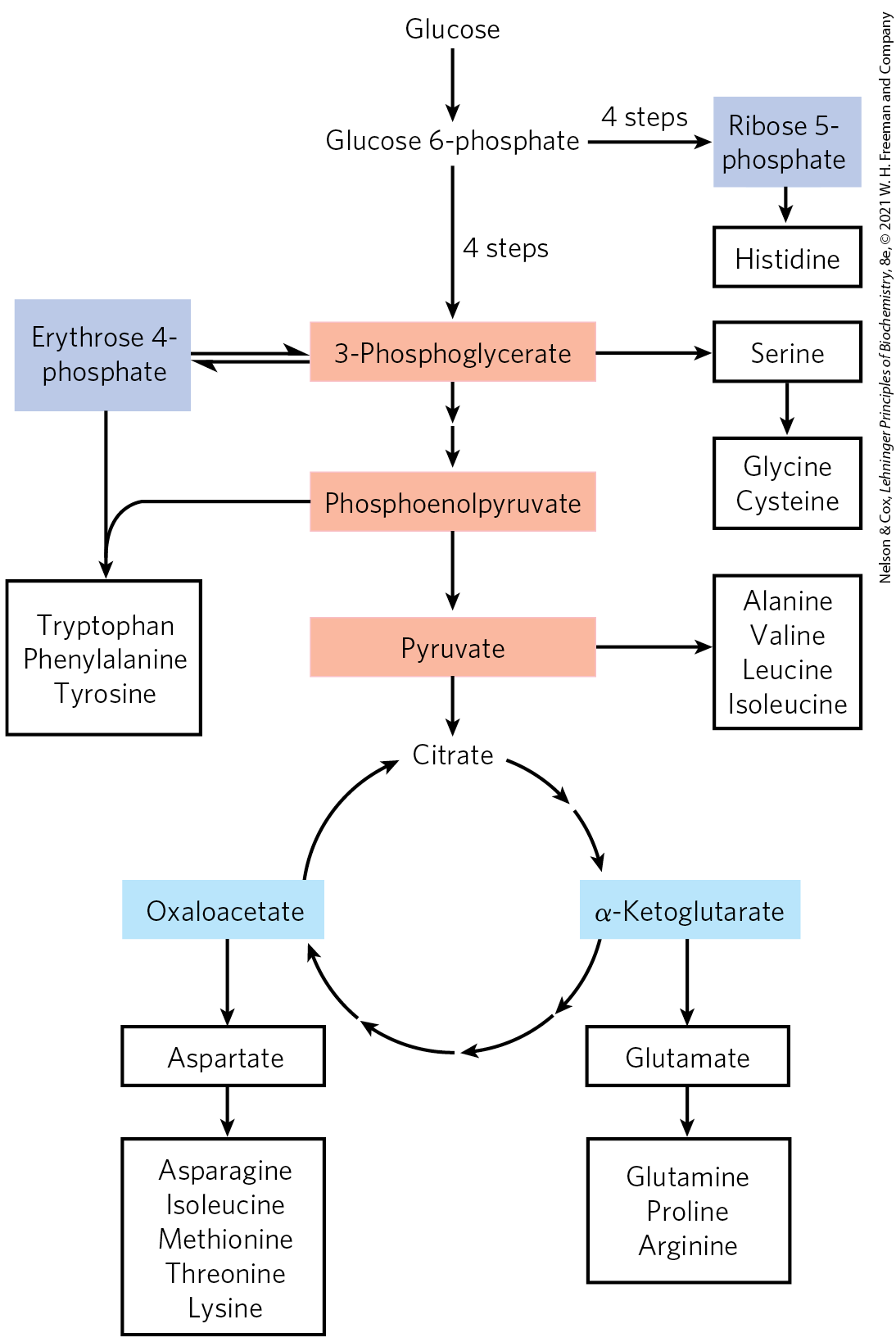
All amino acids are derived from intermediates in glycolysis, the citric acid cycle, or the pentose phosphate pathway (Fig. 22-11). Nitrogen enters these biosynthetic pathways by way of glutamate and glutamine. Some pathways are simple, others are not. Ten of the amino acids are just one or several steps removed from the common metabolite from which they are derived. The biosynthetic pathways for others, such as the aromatic amino acids, are more complex.
FIGURE 22-11 Overview of amino acid biosynthesis. The carbon skeleton precursors derive from three sources: glycolysis (light red), the citric acid cycle (blue), and the pentose phosphate pathway (purple).
Organisms Vary Greatly in Their Ability to Synthesize the 20 Common Amino Acids
Whereas most bacteria and plants can synthesize all 20 amino acids, mammals can synthesize only about half of them — generally those with simple pathways. These are often called nonessential amino acids (see Table 18-1). The label is somewhat misleading, however, because innate biosynthetic pathways often do not provide enough of these amino acids to support optimal growth and health. The remaining amino acids, the essential amino acids, cannot be synthesized by most mammals and must be obtained from food. A few amino acids are conditionally essential in mammals, required at particular stages of development. Unless otherwise indicated, the pathways for the 20 common amino acids presented below are those operative in bacteria.
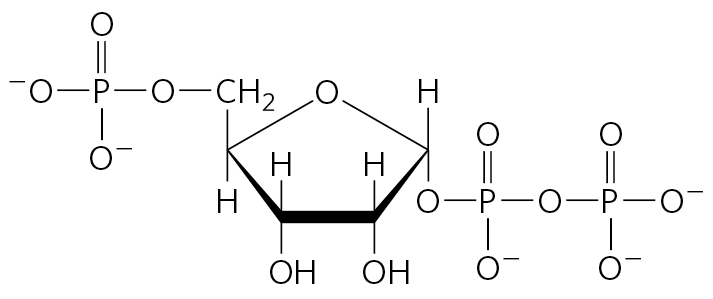
A useful way to organize these biosynthetic pathways is to group them into six families corresponding to their metabolic precursors (Table 22-1), and we use this approach to structure the detailed descriptions that follow. In addition to these six precursors, there is a notable intermediate in several pathways of amino acid and nucleotide synthesis: 5-phosphoribosyl-1-pyrophosphate (PRPP):
TABLE 22-1 Amino Acid Biosynthetic Families, Grouped by Metabolic Precursor
PRPP is synthesized from ribose 5-phosphate derived from the pentose phosphate pathway (see Fig. 14-31), in a reaction catalyzed by ribose phosphate pyrophosphokinase:
This enzyme is allosterically regulated by many of the biomolecules for which PRPP is a precursor.
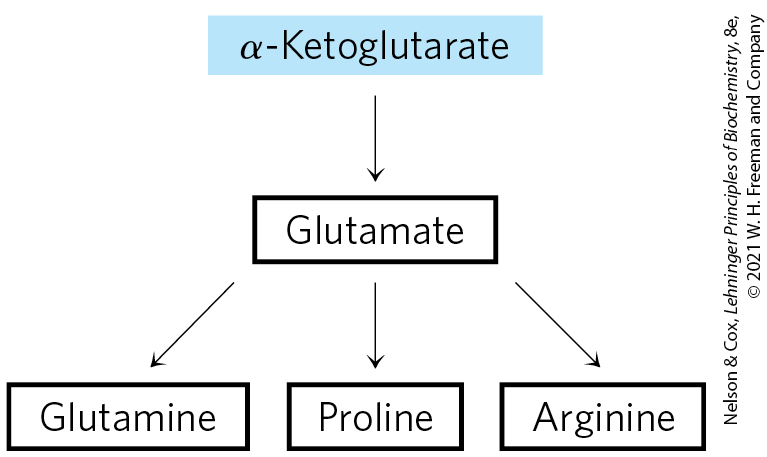
α-Ketoglutarate Gives Rise to Glutamate, Glutamine, Proline, and Arginine
We have already described the biosynthesis of glutamate and glutamine. Proline is a cyclized derivative of glutamate (Fig. 22-12). In the first step of proline synthesis, ATP reacts with the γ-carboxyl group of glutamate to form an acyl phosphate, which is reduced by NADPH or NADH to glutamate γ-semialdehyde. This intermediate undergoes rapid spontaneous cyclization and is then reduced further to yield proline.
FIGURE 22-12 Biosynthesis of proline and arginine from glutamate in bacteria. All five carbon atoms of proline arise from glutamate. In many organisms, glutamate dehydrogenase is unusual in that it uses either NADH or NADPH as a cofactor. The same may be true of other enzymes in these pathways. The γ-semialdehyde in the proline pathway undergoes a rapid, reversible cyclization to -pyrroline-5-carboxylate (P5C), with the equilibrium favoring P5C formation. Cyclization is averted in the ornithine/arginine pathway by acetylation of the α-amino group of glutamate in the first step and removal of the acetyl group after the transamination. This acetyl group is highlighted in yellow. Although some bacteria lack arginase and thus the complete urea cycle, they can synthesize arginine from ornithine in steps that parallel the mammalian urea cycle, with citrulline and argininosuccinate as intermediates (see Fig. 18-10).
Here, and in subsequent figures in this chapter, the reaction arrows indicate the linear path to the final products, without considering the reversibility of individual steps. For example, the step of the pathway leading to arginine that is catalyzed by N-acetylglutamate dehydrogenase is chemically similar to the glyceraldehyde 3-phosphate dehydrogenase reaction in glycolysis (see Fig. 14-7) and is readily reversible.
Arginine is synthesized from glutamate via ornithine and the urea cycle in animals (Chapter 18). In principle, ornithine could also be synthesized from glutamate γ-semialdehyde by transamination, but the spontaneous cyclization of the semialdehyde in the proline pathway precludes a sufficient supply of this intermediate for ornithine synthesis. Bacteria have a de novo biosynthetic pathway for ornithine (and thus arginine) that parallels some steps of the proline pathway but includes two additional steps that avoid the problem of the spontaneous cyclization of glutamate γ-semialdehyde (Fig. 22-12). In the first step, the α-amino group of glutamate is blocked by an acetylation requiring acetyl-CoA; then, after the transamination step, the acetyl group is removed to yield ornithine.
The pathways to proline and arginine are somewhat different in mammals. Proline can be synthesized by the pathway shown in Figure 22-12, but it is also formed from arginine obtained from dietary or tissue protein. Arginase, a urea cycle enzyme, converts arginine to ornithine and urea (see Figs. 18-10, 18-26). The ornithine is converted to glutamate γ-semialdehyde by the enzyme ornithine γ-aminotransferase (Fig. 22-13). The semialdehyde cyclizes to -pyrroline-5-carboxylate, which is then converted to proline (Fig. 22-12). The pathway for arginine synthesis shown in Figure 22-12 is absent in mammals. When arginine from dietary intake or protein turnover is insufficient for protein synthesis, the ornithine δ-aminotransferase reaction operates in the direction of ornithine formation. Ornithine is then converted to citrulline and arginine in the urea cycle.
FIGURE 22-13 Ornithine δ-aminotransferase reaction: a step in the mammalian pathway to proline. This enzyme is found in the mitochondrial matrix of most tissues. Although the equilibrium favors P5C formation, the reverse reaction is the only mammalian pathway for synthesis of ornithine (and thus arginine) when arginine levels are insufficient for protein synthesis.
Serine, Glycine, and Cysteine Are Derived from 3-Phosphoglycerate
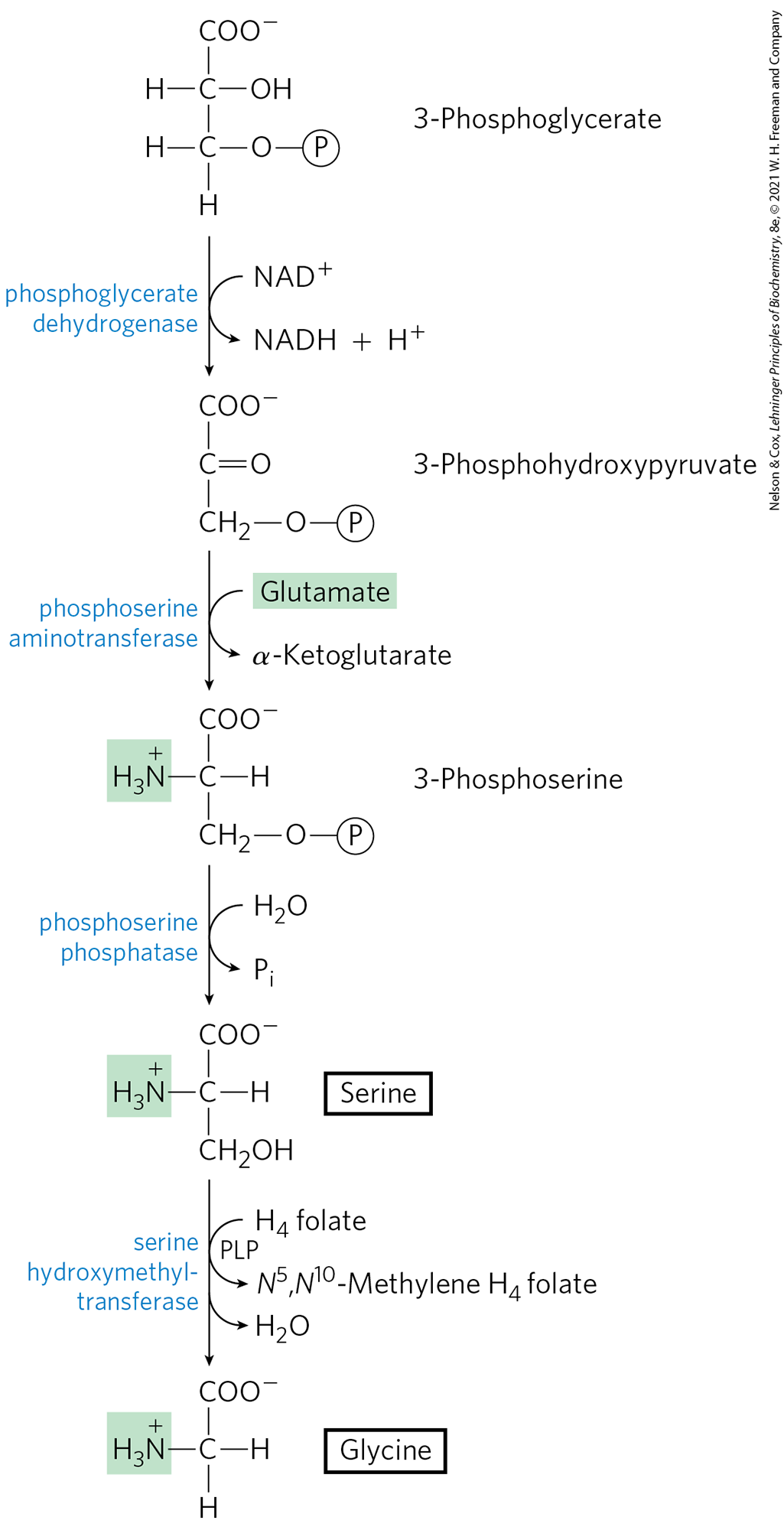
The major pathway for the formation of serine is the same in all organisms (Fig. 22-14). In the first step, the hydroxyl group of 3-phosphoglycerate is oxidized by a dehydrogenase (using ) to yield 3-phosphohydroxypyruvate. Transamination from glutamate yields 3-phosphoserine, which is hydrolyzed to free serine by phosphoserine phosphatase.
FIGURE 22-14 Biosynthesis of serine from 3-phosphoglycerate and of glycine from serine in all organisms. As indicated in the text, this is only one of multiple pathways to synthesize glycine.
Serine (three carbons) is the precursor of glycine (two carbons) through removal of a carbon atom by serine hydroxymethyltransferase (Fig. 22-14). Tetrahydrofolate accepts the β carbon (C-3) of serine, which forms a methylene bridge between N-5 and N-10 to yield -methylenetetrahydrofolate (see Fig. 18-17). The overall reaction, which is reversible, also requires pyridoxal phosphate. In the liver of vertebrates, glycine can be made by another route: the reverse of the reaction shown in Figure 18-20c, catalyzed by glycine synthase (also called glycine cleavage enzyme):
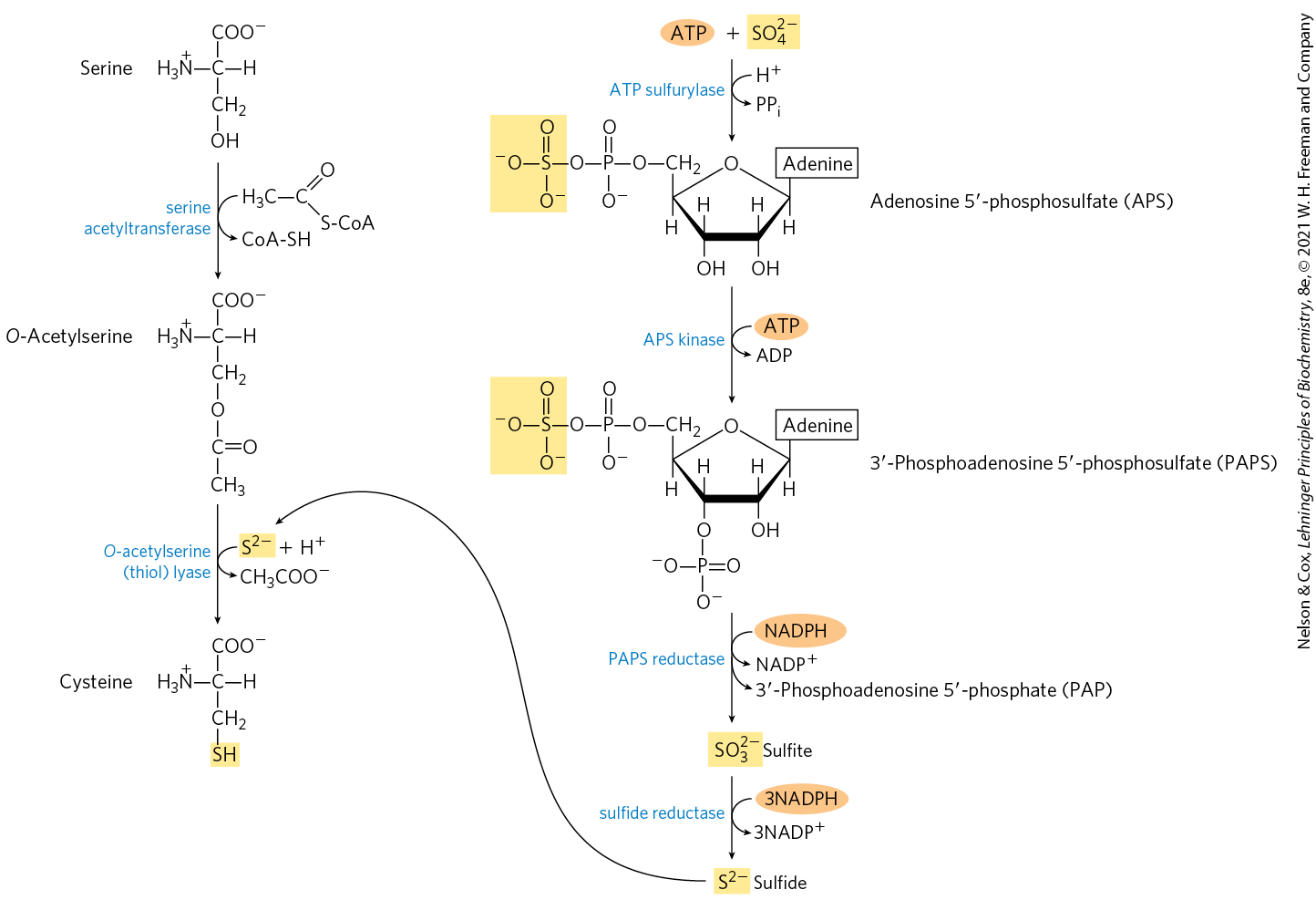
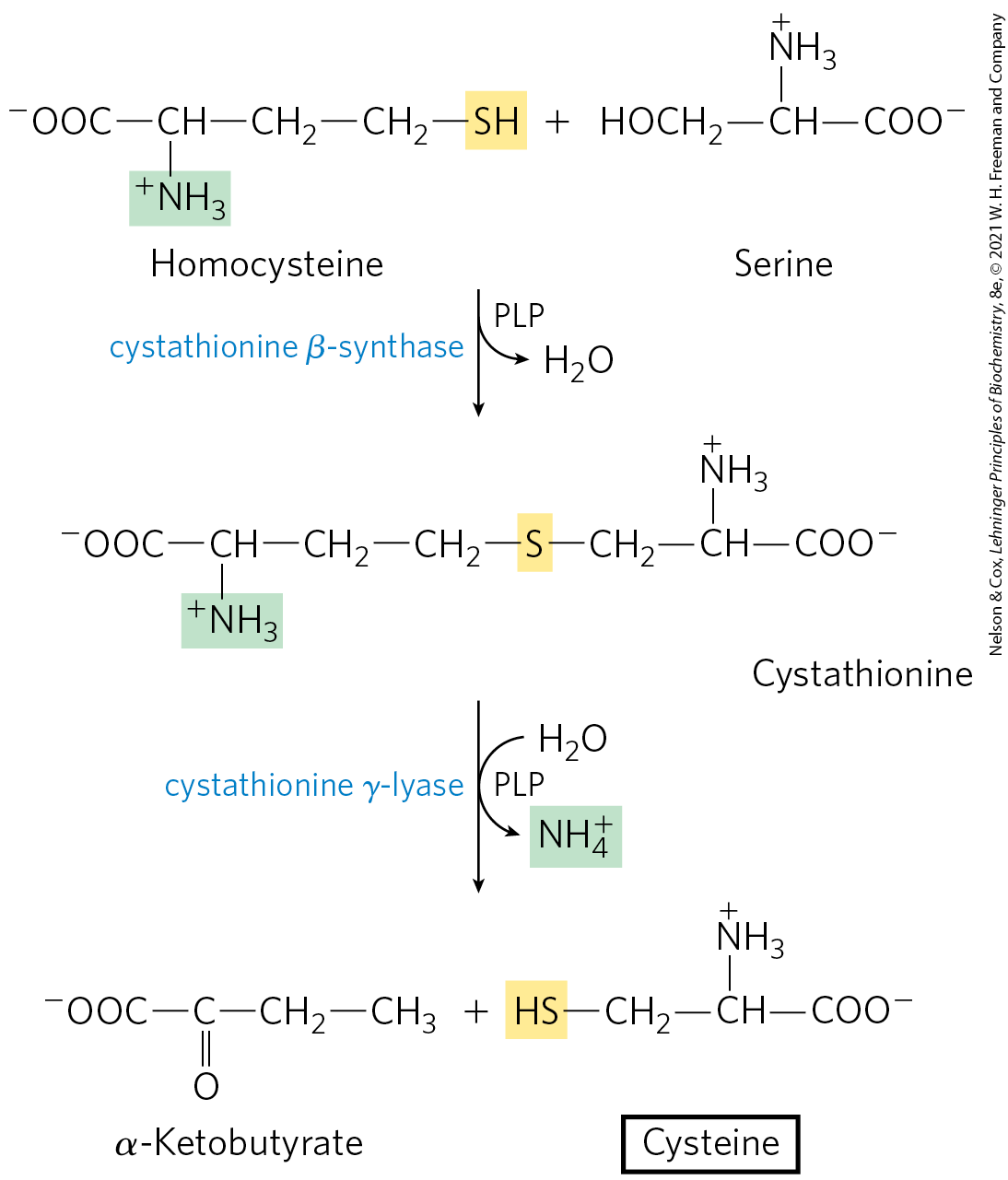
Plants and bacteria produce the reduced sulfur required for the synthesis of cysteine (and methionine, described later) from environmental sulfates; the pathway is shown on the right side of Figure 22-15. Sulfate is activated in two steps to produce -phosphoadenosine -phosphosulfate (PAPS), which undergoes an eight-electron reduction to sulfide. The sulfide is then used in the formation of cysteine from serine in a two-step pathway. Mammals synthesize cysteine from two amino acids: methionine furnishes the sulfur atom, and serine furnishes the carbon skeleton. Methionine is first converted to S-adenosylmethionine (see Fig. 18-18), which can lose its methyl group to any of a number of acceptors to form S-adenosylhomocysteine (adoHcy). This demethylated product is hydrolyzed to free homocysteine, which undergoes a reaction with serine, catalyzed by cystathionine β-synthase, to yield cystathionine (Fig. 22-16). Finally, cystathionine δ-lyase, a PLP-requiring enzyme, catalyzes removal of ammonia and cleavage of cystathionine to yield free cysteine.
FIGURE 22-15 Biosynthesis of cysteine from serine in bacteria and plants. The origin of reduced sulfur is shown in the pathway on the right.
FIGURE 22-16 Biosynthesis of cysteine from homocysteine and serine in mammals. The homocysteine is formed from methionine.
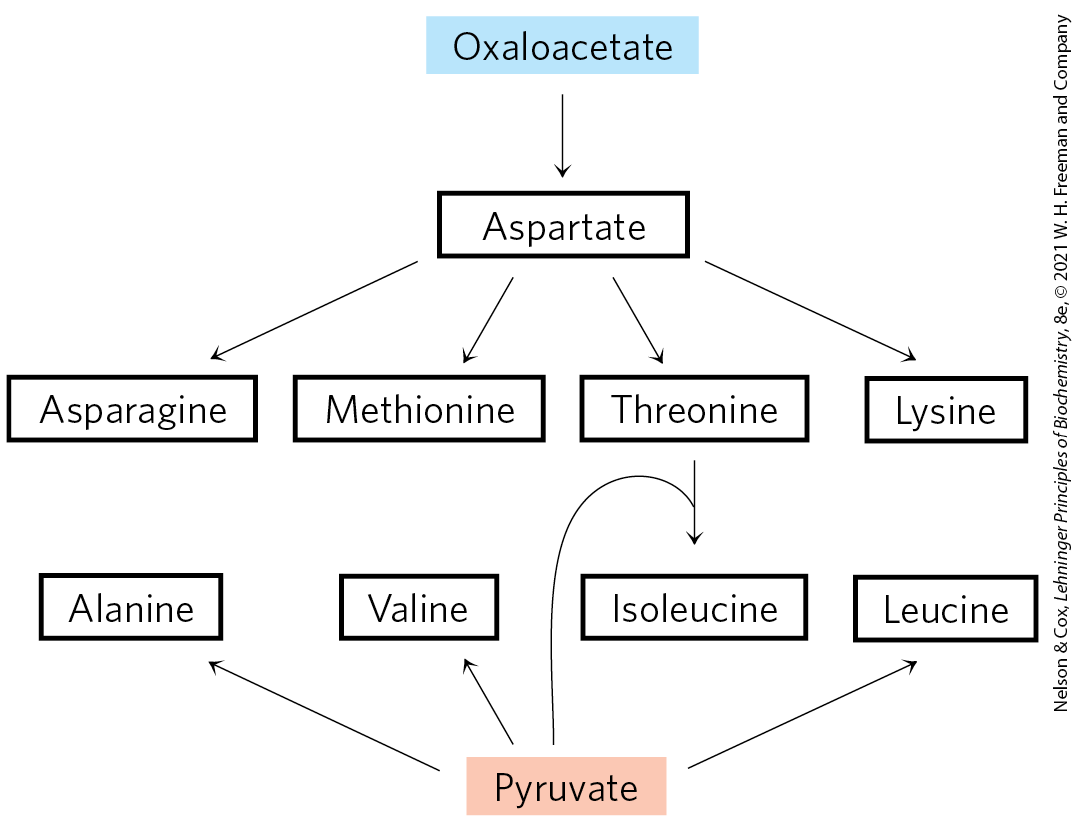
Three Nonessential and Six Essential Amino Acids Are Synthesized from Oxaloacetate and Pyruvate
Alanine and aspartate are synthesized from pyruvate and oxaloacetate, respectively, by transamination from glutamate. Asparagine is synthesized by amidation of aspartate, catalyzed by the enzyme asparagine synthetase. The is donated by glutamine. These are nonessential amino acids, and their simple biosynthetic pathways occur in all organisms.
The malignant lymphocytes present in childhood acute lymphoblastic leukemia (ALL) produce little or no asparagine synthetase, and they are thus sensitive to asparagine depletion. The chemotherapy for ALL is administered together with an l-asparaginase derived from bacteria, with the enzyme functioning to reduce serum asparagine. The combined treatment results in a greater than 95% remission rate in cases of childhood ALL (l-asparaginase treatment alone produces remission in 40% to 60% of cases). However, the asparaginase treatment has some deleterious side effects, and about 10% of patients who achieve remission eventually suffer relapse, with tumors resistant to drug therapy. Researchers are now developing inhibitors of human asparagine synthetase to augment these therapies for childhood ALL.
Methionine, threonine, lysine, isoleucine, valine, and leucine are essential amino acids; humans cannot synthesize them. Their biosynthetic pathways in bacteria are complex and interconnected (Fig. 22-17). In some cases, the pathways in bacteria, fungi, and plants differ significantly.
FIGURE 22-17 Biosynthesis of six essential amino acids from oxaloacetate and pyruvate in bacteria: methionine, threonine, lysine, isoleucine, valine, and leucine. Some of the most complex pathways for amino acid biosynthesis are found here. Pathways are abbreviated to emphasize precursors and pathway products.
Aspartate gives rise to methionine, threonine, and lysine. Branch points occur at aspartate β-semialdehyde, an intermediate in all three pathways, and at homoserine, a precursor of threonine and methionine. Threonine, in turn, is one of the precursors of isoleucine. The valine and isoleucine pathways share four enzymes (Fig. 22-17). Pyruvate gives rise to valine and isoleucine in pathways that begin with condensation of two carbons of pyruvate (in the form of hydroxyethyl thiamine pyrophosphate; see Fig. 14-13b) with another molecule of pyruvate (the valine path) or with α-ketobutyrate (the isoleucine path). The α-ketobutyrate is derived from threonine in a reaction that requires pyridoxal phosphate. An intermediate in the valine pathway, α-ketoisovalerate, is the starting point for a four-step branch pathway leading to leucine.
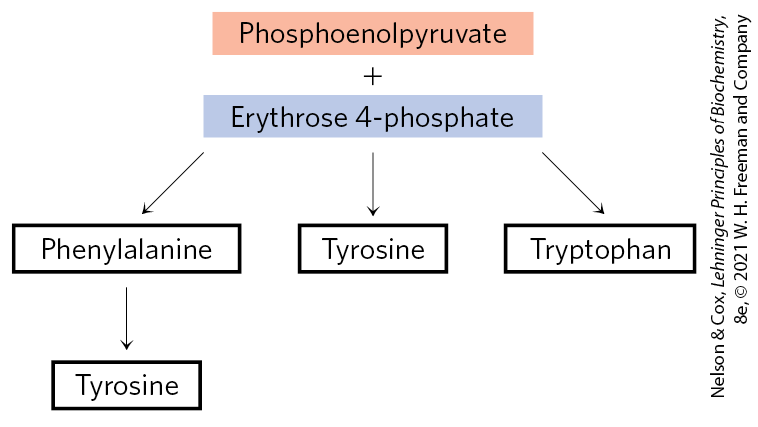
Chorismate Is a Key Intermediate in the Synthesis of Tryptophan, Phenylalanine, and Tyrosine
The three amino acid side chains that contain aromatic rings, tryptophan, phenylalanine, and tyrosine, present a special chemical problem for biosynthesis. Aromatic rings are not readily available in the environment, even though the benzene ring is very stable. The branched pathway to tryptophan, phenylalanine, and tyrosine, occurring in bacteria, fungi, and plants, is the main biological route of aromatic ring formation. It proceeds through ring closure of an aliphatic precursor followed by stepwise addition of double bonds. The first four steps produce shikimate, a seven-carbon molecule derived from erythrose 4-phosphate and phosphoenolpyruvate (Fig. 22-18). Shikimate is converted to chorismate in three steps that include the addition of three more carbons from another molecule of phosphoenolpyruvate. Chorismate is the first branch point of the pathway, with one branch leading to tryptophan, the other to phenylalanine and tyrosine.
FIGURE 22-18 Biosynthesis of chorismate, an intermediate in the synthesis of aromatic amino acids in bacteria and plants. All carbons are derived from either erythrose 4-phosphate (light purple) or phosphoenolpyruvate (light red). Note that the required as a cofactor in step is released unchanged; it may be transiently reduced to NADH during the reaction, with formation of an oxidized reaction intermediate. Step is competitively inhibited by glyphosate the active ingredient in the widely used herbicide Roundup. The herbicide is relatively nontoxic to mammals, which lack this biosynthetic pathway. The intermediates quinate and shikimate are named after the plants in which they have been found to accumulate.
In the tryptophan branch (Fig. 22-19), chorismate is converted to anthranilate in a reaction in which glutamine donates the nitrogen that will become part of the indole ring. Anthranilate then condenses with PRPP. The indole ring of tryptophan is derived from the ring carbons and amino group of anthranilate plus two carbons derived from PRPP. The final reaction in the sequence is catalyzed by tryptophan synthase. This enzyme has an subunit structure and can be dissociated into two α subunits and a unit that catalyze different parts of the overall reaction:
FIGURE 22-19 Biosynthesis of tryptophan from chorismate in bacteria and plants. In E. coli, enzymes catalyzing steps and are subunits of a single complex.
The second part of the reaction requires pyridoxal phosphate (Fig. 22-20). Indole formed in the first part is not released by the enzyme, but instead moves through a channel from the α-subunit active site to one of the β-subunit active sites, where it condenses with a Schiff base intermediate derived from serine and PLP. Intermediate channeling of this type may be a feature of the entire pathway from chorismate to tryptophan. Enzyme active sites catalyzing different steps (sometimes not sequential steps) of the pathway to tryptophan are found on single polypeptides in some species of fungi and bacteria, but they are separate proteins in other species. In addition, the activity of some of these enzymes requires a noncovalent association with other enzymes of the pathway. These observations suggest that all the pathway enzymes are components of a large, multienzyme complex, a metabolon, in both bacteria and eukaryotes. Such complexes are generally not preserved intact when the enzymes are isolated using traditional biochemical methods (see Section 16.4).
MECHANISM FIGURE 22-20 Tryptophan synthase reaction. (a) This enzyme catalyzes a multistep reaction with several types of chemical rearrangements. The PLP-facilitated transformations occur at the β carbon (C-3) of the amino acid, as opposed to the α-carbon reactions described in Figure 18-6. The β carbon of serine is attached to the indole ring system. (b) Indole generated on the α subunit (white) moves through a tunnel to the β subunit (blue), where it condenses with the serine moiety. [(b) Data from PDB ID 1KFJ, V. Kulik et al., J. Mol. Biol. 324:677, 2002.]
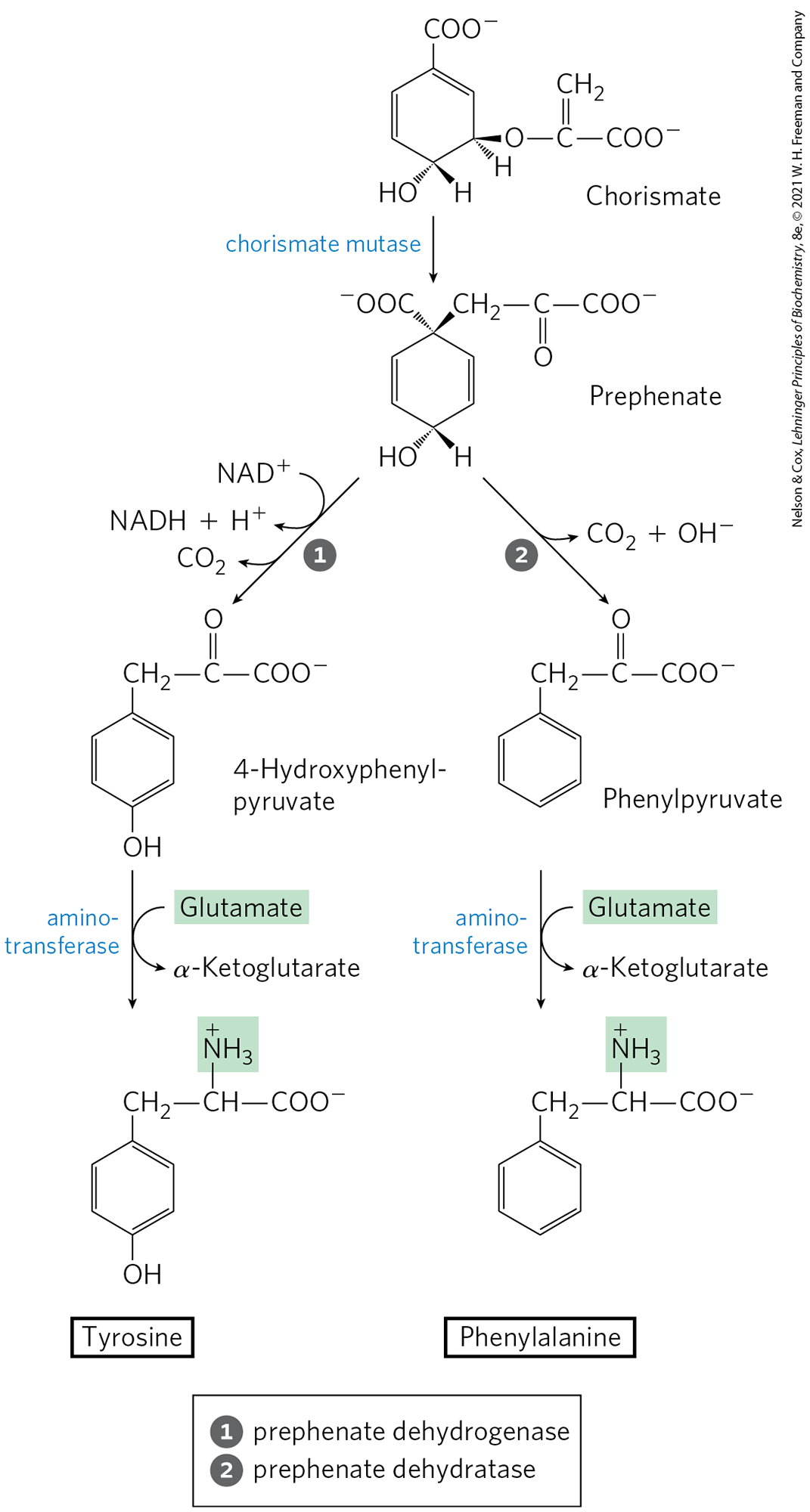
In plants and bacteria, phenylalanine and tyrosine are synthesized from chorismate in pathways much less complex than the tryptophan pathway. The common intermediate is prephenate (Fig. 22-21). The final step in both cases is transamination with glutamate.
FIGURE 22-21 Biosynthesis of phenylalanine and tyrosine from chorismate in bacteria and plants. Conversion of chorismate to prephenate is a rare biological example of a Claisen rearrangement.
Animals can produce tyrosine directly from phenylalanine through hydroxylation at C-4 of the phenyl group by phenylalanine hydroxylase; this enzyme also participates in the degradation of phenylalanine (see Figs. 18-23, 18-24). Tyrosine is considered a conditionally essential amino acid, or as nonessential insofar as it can be synthesized from the essential amino acid phenylalanine.
Histidine Biosynthesis Uses Precursors of Purine Biosynthesis
The pathway to histidine in all plants and bacteria differs in several respects from other amino acid biosynthetic pathways. Histidine is derived from three precursors (Fig. 22-22): PRPP contributes five carbons, the purine ring of ATP contributes a nitrogen and a carbon, and glutamine supplies the second ring nitrogen. The key steps are condensation of ATP and PRPP, in which N-1 of the purine ring is linked to the activated C-1 of the ribose of PRPP (step in Fig. 22-22); purine ring opening that ultimately leaves N-1 and C-2 of adenine linked to the ribose (step ); and formation of the imidazole ring, a reaction in which glutamine donates a nitrogen (step ). The use of ATP as a metabolite rather than a high-energy cofactor is unusual — but not wasteful, because it dovetails with the purine biosynthetic pathway. The remnant of ATP that is released after the transfer of N-1 and C-2 is 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR), an intermediate of purine biosynthesis (see Fig. 22-35) that is rapidly recycled to ATP.
FIGURE 22-22 Biosynthesis of histidine in bacteria and plants. Atoms derived from PRPP and ATP are shaded light red and blue, respectively. Two of the histidine nitrogens are derived from glutamine and glutamate (green). Note that the derivative of ATP remaining after step (AICAR) is an intermediate in purine biosynthesis (see Fig. 22-35, step ), so ATP is rapidly regenerated.
Amino Acid Biosynthesis Is under Allosteric Regulation
As detailed in Chapter 13, the control of flux through a metabolic pathway often reflects the activity of multiple enzymes in that pathway. In the case of amino acid synthesis, regulation often takes place in part through feedback inhibition of the first reaction by the end product of the pathway. This first reaction is often catalyzed by an allosteric enzyme that plays an important role in the overall control of flux through that pathway. As an example, Figure 22-23 shows the allosteric regulation of isoleucine synthesis from threonine. The end product, isoleucine, is an allosteric inhibitor of the first reaction in the sequence. In bacteria, such allosteric modulation of amino acid synthesis contributes to the minute-to-minute adjustment of pathway activity to cellular needs.
FIGURE 22-23 Allosteric regulation of isoleucine biosynthesis. The first reaction in the pathway from threonine to isoleucine is inhibited by the end product, isoleucine. This was one of the first examples of allosteric feedback inhibition to be discovered.
Allosteric regulation of an individual enzyme can be considerably more complex. An example is the remarkable set of allosteric controls exerted on glutamine synthetase of E. coli (Fig. 22-8). Six products derived from glutamine serve as negative feedback modulators of the enzyme, and the overall effects of these and other modulators are more than additive. Such regulation is called concerted inhibition.
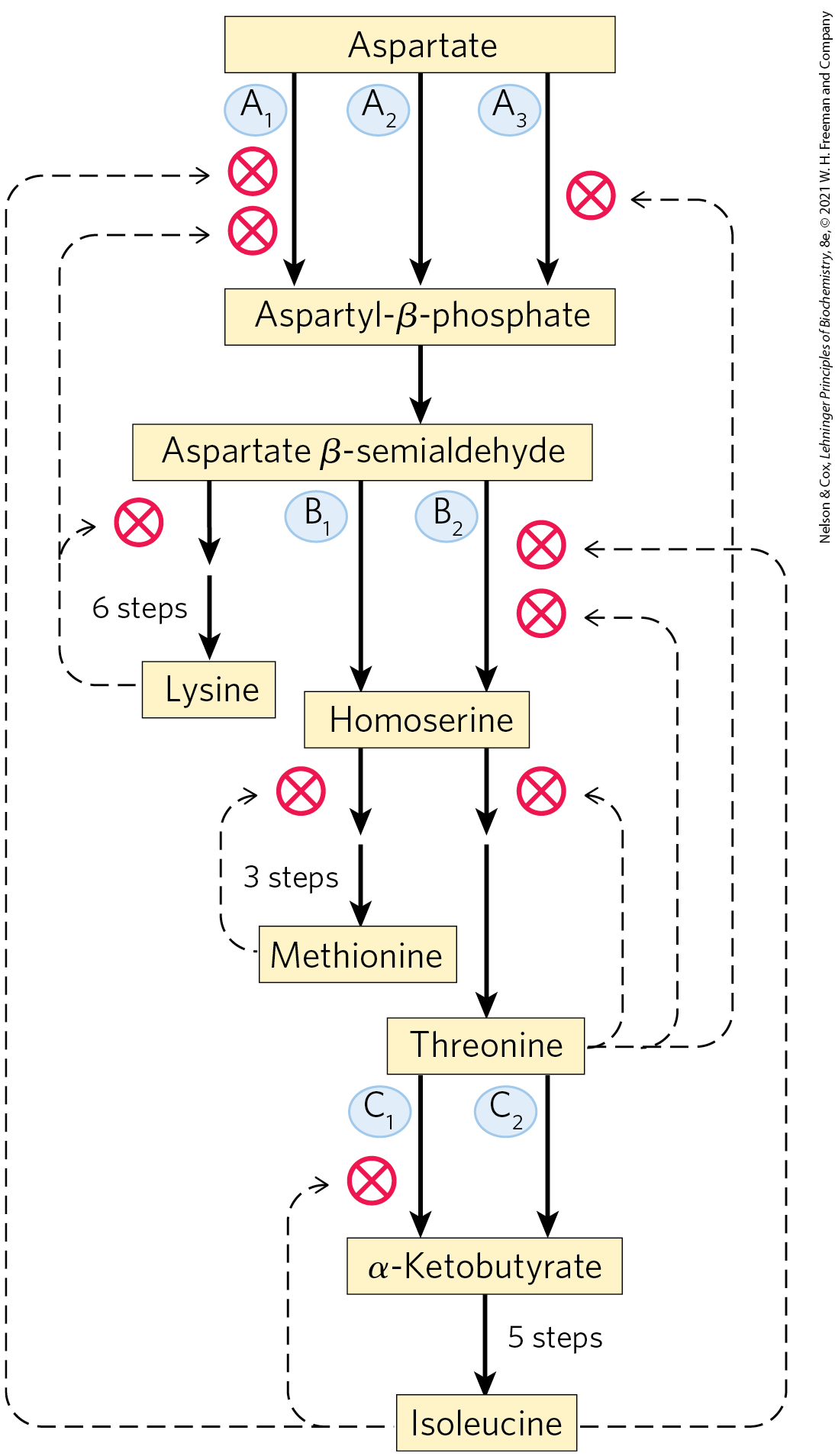
Additional mechanisms contribute to the regulation of the amino acid biosynthetic pathways. Because the 20 common amino acids must be made in the correct proportions for protein synthesis, cells have developed ways not only of controlling the rate of synthesis of individual amino acids but also of coordinating their formation. Such coordination is especially well developed in fast-growing bacterial cells. Figure 22-24 shows how E. coli cells coordinate the synthesis of lysine, methionine, threonine, and isoleucine, all made from aspartate. Several important types of inhibition patterns are evident. The step from aspartate to aspartyl-β-phosphate is catalyzed by three isozymes, each independently controlled by different modulators. This enzyme multiplicity prevents one biosynthetic end product from shutting down key steps in a pathway when other products of the same pathway are required. The steps from aspartate β-semialdehyde to homoserine and from threonine to α-ketobutyrate (detailed in Fig. 22-17) are also catalyzed by dual, independently controlled isozymes. One isozyme for the conversion of aspartate to aspartyl-β-phosphate is allosterically inhibited by two different modulators, lysine and isoleucine, whose action is more than additive — another example of concerted inhibition. The sequence from aspartate to isoleucine undergoes multiple, overlapping negative feedback inhibitions; for example, isoleucine inhibits the conversion of threonine to α-ketobutyrate (as described above), and threonine inhibits its own formation at three points: from homoserine, from aspartate β-semialdehyde, and from aspartate (see Fig. 22-17). This overall regulatory mechanism is called sequential feedback inhibition.
FIGURE 22-24 Interlocking regulatory mechanisms in the biosynthesis of several amino acids derived from aspartate in E. coli. Three enzymes (A, B, C) have either two or three isozyme forms, indicated by numerical subscripts. In each case, one isozyme has no allosteric regulation; these isozymes are regulated by changes in the amount of enzyme synthesized. Synthesis of isozymes and is repressed when methionine levels are high, and synthesis of isozyme is repressed when isoleucine levels are high. Enzyme A is aspartokinase; B, homoserine dehydrogenase; C, threonine dehydratase.
Similar patterns are evident in the pathways leading to the aromatic amino acids. The first step of the early pathway to the common intermediate chorismate is catalyzed by the enzyme 2-keto-3-deoxy-d1-arabinoheptulosonate 7-phosphate (DAHP) synthase ( in Fig. 22-18). Most microorganisms and plants have three DAHP synthase isozymes. One is allosterically inhibited (feedback inhibition) by phenylalanine, another by tyrosine, and the third by tryptophan. This scheme helps the overall pathway to respond to cellular requirements for one or more of the aromatic amino acids. Additional regulation takes place after the pathway branches at chorismate. For example, the enzymes catalyzing the first two steps of the tryptophan branch are subject to allosteric inhibition by tryptophan.
SUMMARY 22.2 Biosynthesis of Amino Acids
Plants and bacteria synthesize all 20 common amino acids. Mammals can synthesize about half; the others are required in the diet (essential or conditionally essential amino acids).
Glutamate is formed by reductive amination of α-ketoglutarate and serves as the precursor of glutamine, proline, and arginine.
The carbon chain of serine is derived from 3-phosphoglycerate. Serine is a precursor of glycine; the β-carbon atom of serine is transferred to tetrahydrofolate. In microorganisms, cysteine is produced from serine and from sulfide produced by the reduction of environmental sulfate. Mammals produce cysteine from methionine and serine by a series of reactions requiring S-adenosylmethionine and cystathionine.
Alanine and aspartate (and thus asparagine) are formed from pyruvate and oxaloacetate, respectively, by transamination. Pyruvate and oxaloacetate also give rise to methionine, threonine, lysine, valine, isoleucine, and leucine via longer pathways.
The aromatic amino acids (phenylalanine, tyrosine, and tryptophan) form by a pathway in which chorismate occupies a key branch point. Tyrosine can also be formed by hydroxylation of phenylalanine (and thus is considered conditionally essential). Phosphoribosyl pyrophosphate is a precursor of tryptophan and histidine.
The pathway to histidine is interconnected with the purine synthetic pathway.
The amino acid biosynthetic pathways are subject to allosteric end-product inhibition; the regulatory enzyme is usually the first in the sequence. Regulation of the various synthetic pathways is coordinated.
 In the first step of proline synthesis, ATP reacts with the γ-carboxyl group of glutamate to form an acyl phosphate, which is reduced by NADPH or NADH to glutamate γ-semialdehyde. This intermediate undergoes rapid spontaneous cyclization and is then reduced further to yield proline.
In the first step of proline synthesis, ATP reacts with the γ-carboxyl group of glutamate to form an acyl phosphate, which is reduced by NADPH or NADH to glutamate γ-semialdehyde. This intermediate undergoes rapid spontaneous cyclization and is then reduced further to yield proline.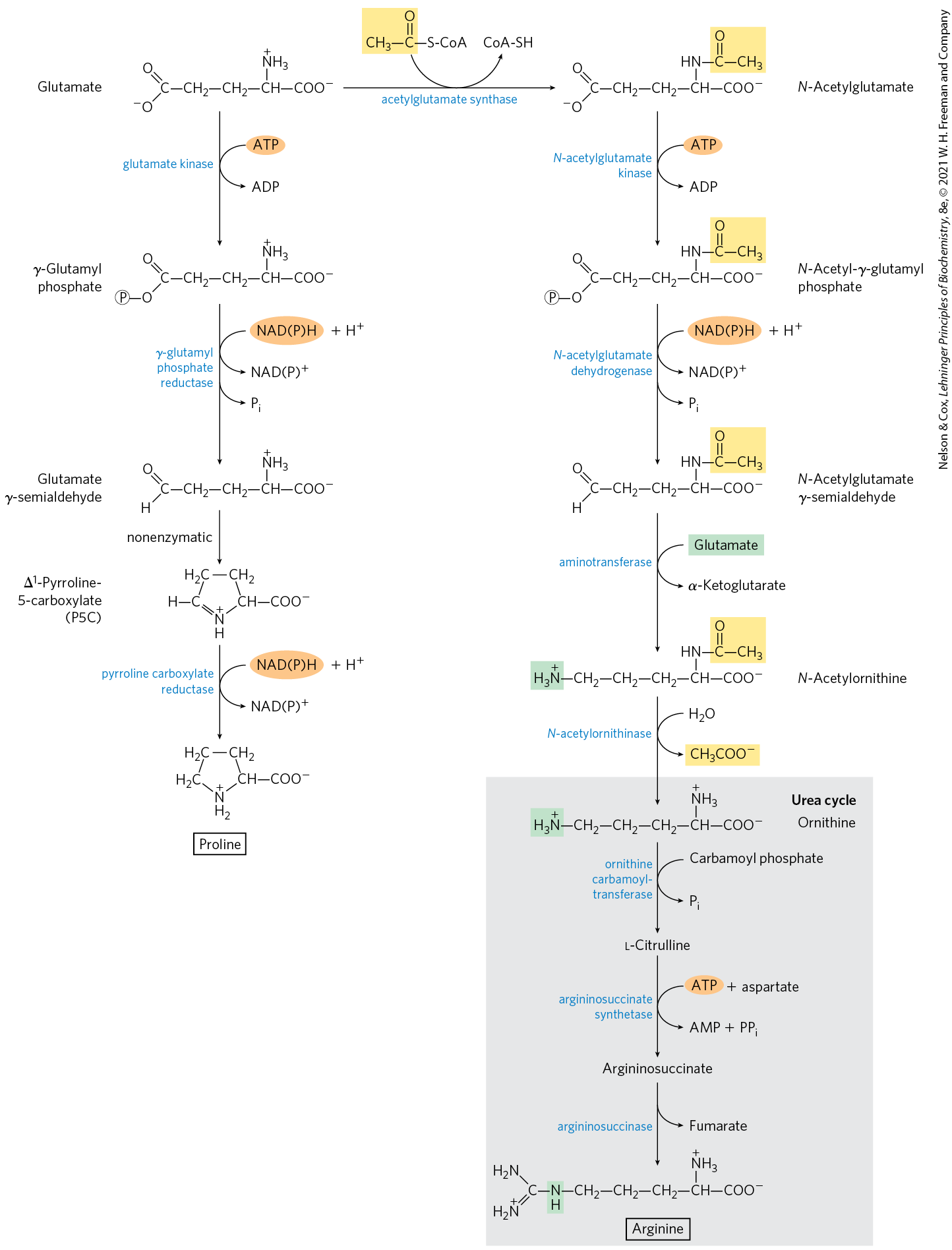
 When arginine from dietary intake or protein turnover is insufficient for protein synthesis, the ornithine δ-aminotransferase reaction operates in the direction of ornithine formation. Ornithine is then converted to citrulline and arginine in the urea cycle.
When arginine from dietary intake or protein turnover is insufficient for protein synthesis, the ornithine δ-aminotransferase reaction operates in the direction of ornithine formation. Ornithine is then converted to citrulline and arginine in the urea cycle.
 The malignant lymphocytes present in childhood acute lymphoblastic leukemia (ALL) produce little or no asparagine synthetase, and they are thus sensitive to asparagine depletion. The chemotherapy for ALL is administered together with an
The malignant lymphocytes present in childhood acute lymphoblastic leukemia (ALL) produce little or no asparagine synthetase, and they are thus sensitive to asparagine depletion. The chemotherapy for ALL is administered together with an 

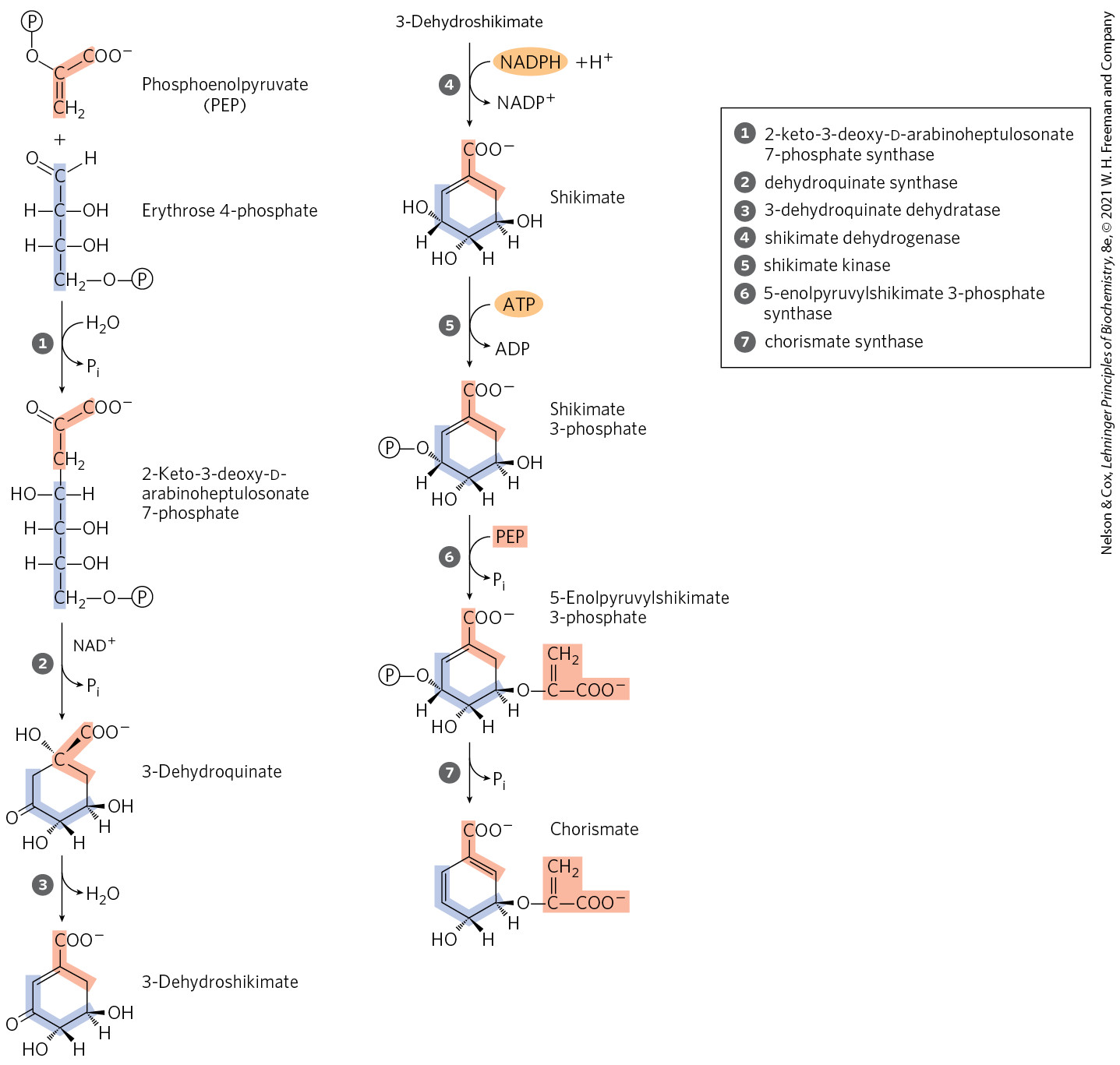
 is released unchanged; it may be transiently reduced to NADH during the reaction, with formation of an oxidized reaction intermediate. Step
is released unchanged; it may be transiently reduced to NADH during the reaction, with formation of an oxidized reaction intermediate. Step  is competitively inhibited by glyphosate the active ingredient in the widely used herbicide Roundup. The herbicide is relatively nontoxic to mammals, which lack this biosynthetic pathway. The intermediates quinate and shikimate are named after the plants in which they have been found to accumulate.
is competitively inhibited by glyphosate the active ingredient in the widely used herbicide Roundup. The herbicide is relatively nontoxic to mammals, which lack this biosynthetic pathway. The intermediates quinate and shikimate are named after the plants in which they have been found to accumulate.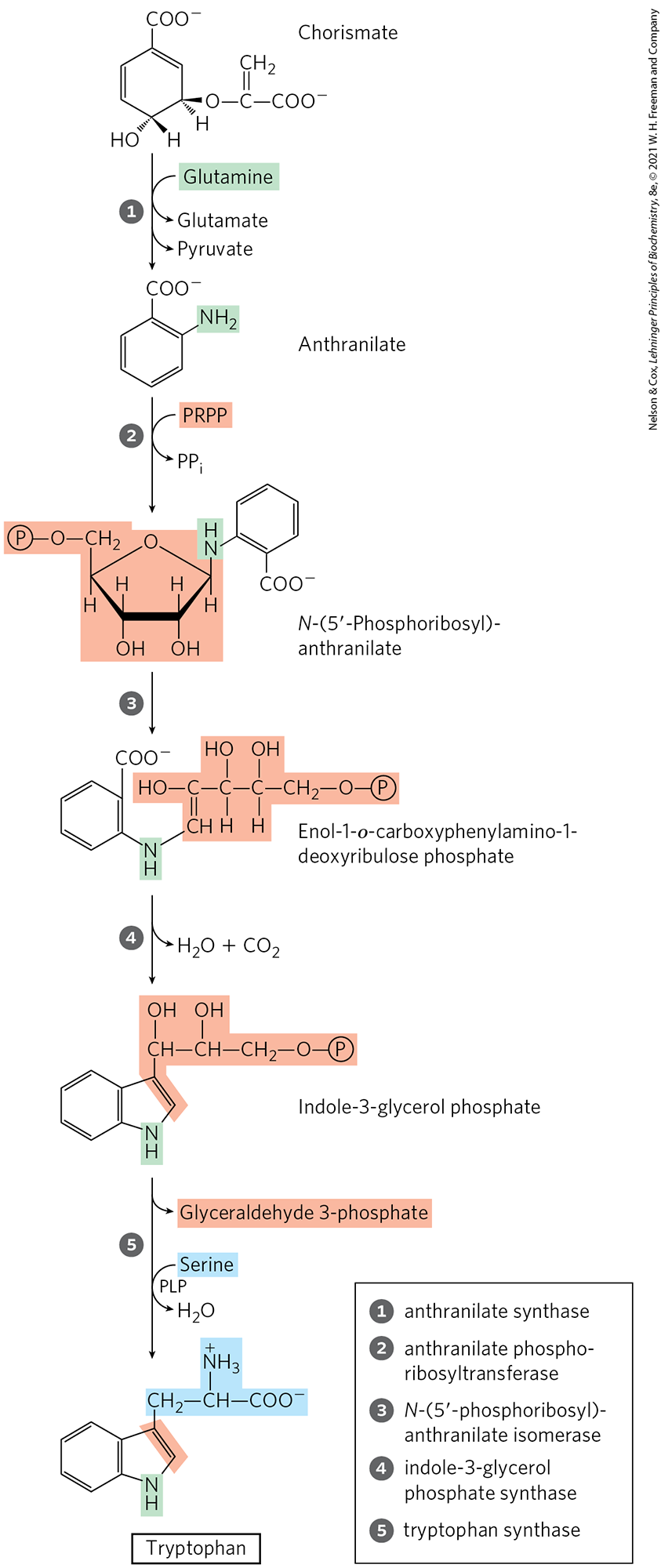
 and
and 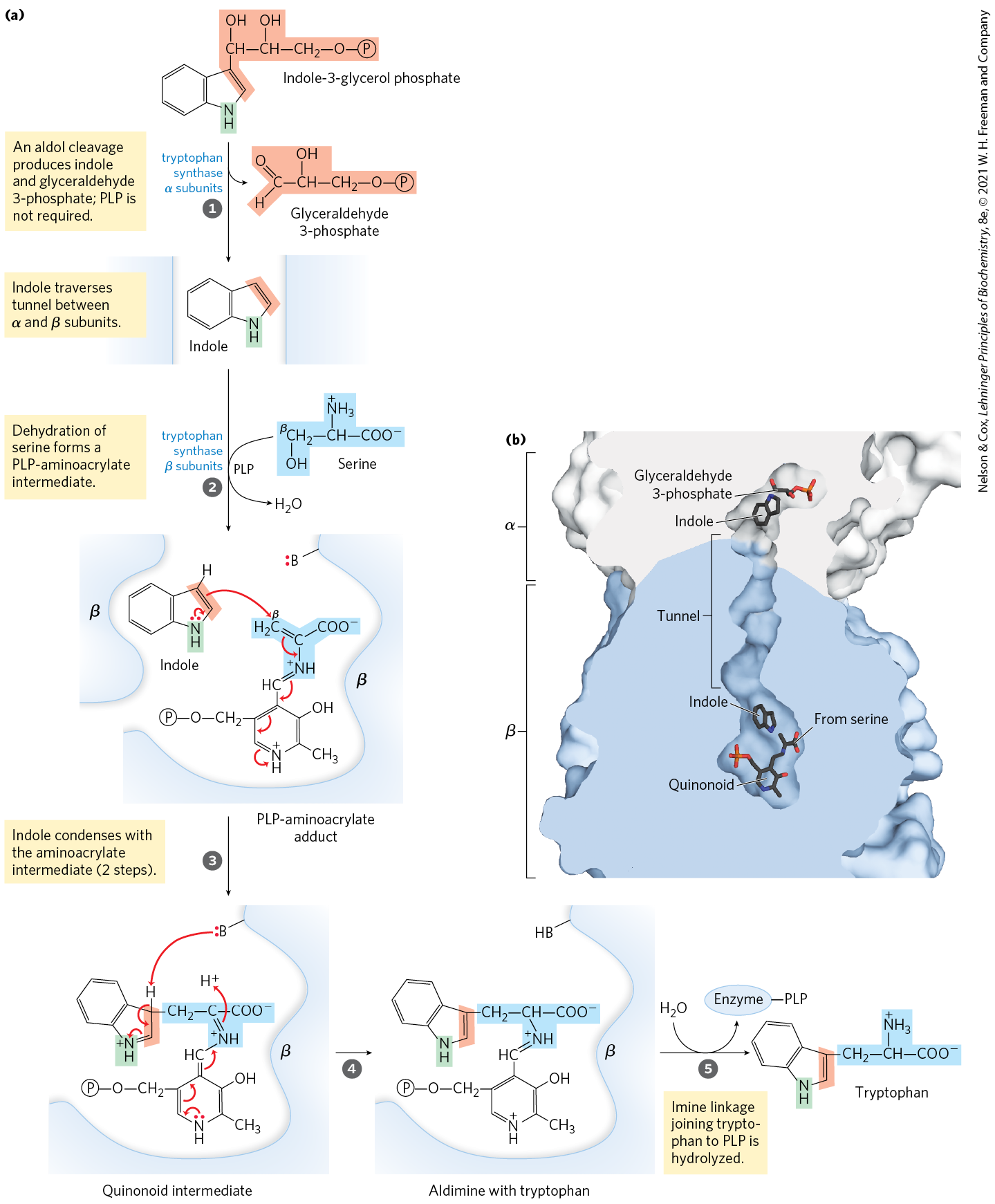
 ); and formation of the imidazole ring, a reaction in which glutamine donates a nitrogen (step
); and formation of the imidazole ring, a reaction in which glutamine donates a nitrogen (step  ). The use of ATP as a metabolite rather than a high-energy cofactor is unusual — but not wasteful, because it dovetails with the purine biosynthetic pathway. The remnant of ATP that is released after the transfer of N-1 and C-2 is 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR), an intermediate of purine biosynthesis (see
). The use of ATP as a metabolite rather than a high-energy cofactor is unusual — but not wasteful, because it dovetails with the purine biosynthetic pathway. The remnant of ATP that is released after the transfer of N-1 and C-2 is 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR), an intermediate of purine biosynthesis (see 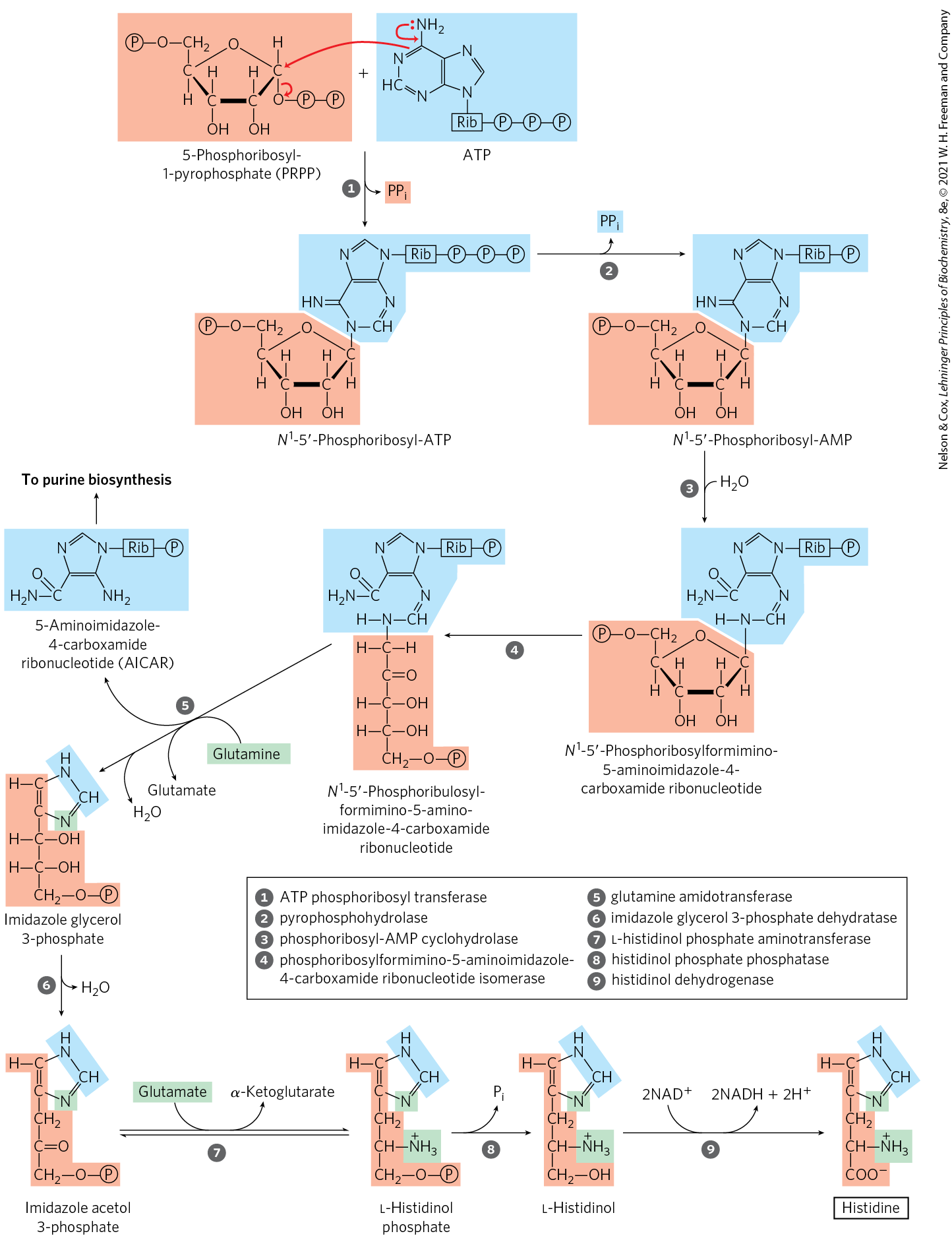
 ), so ATP is rapidly regenerated.
), so ATP is rapidly regenerated. In the case of amino acid synthesis, regulation often takes place in part through feedback inhibition of the first reaction by the end product of the pathway. This first reaction is often catalyzed by an allosteric enzyme that plays an important role in the overall control of flux through that pathway. As an example,
In the case of amino acid synthesis, regulation often takes place in part through feedback inhibition of the first reaction by the end product of the pathway. This first reaction is often catalyzed by an allosteric enzyme that plays an important role in the overall control of flux through that pathway. As an example, 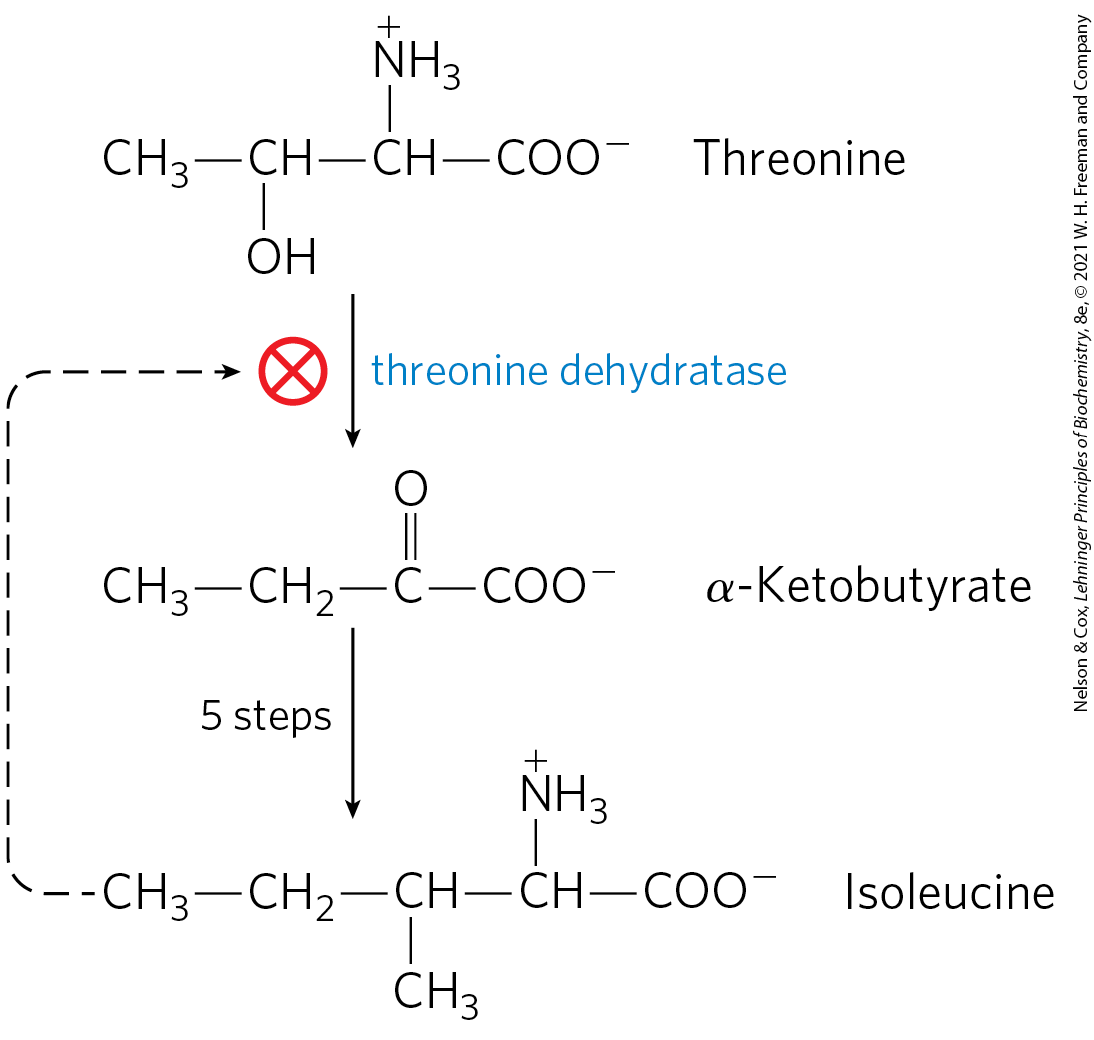
 Plants and bacteria synthesize all 20 common amino acids. Mammals can synthesize about half; the others are required in the diet (essential or conditionally essential amino acids).
Plants and bacteria synthesize all 20 common amino acids. Mammals can synthesize about half; the others are required in the diet (essential or conditionally essential amino acids).