The eight-step cyclic process for oxidation of simple two-carbon acetyl groups to may seem unnecessarily complex and not in keeping with the biological principle of maximum economy. Remember, though, that the role of the citric acid cycle is not confined to the oxidation of acetate from carbohydrates, fatty acids, or amino acids. The cycle also accepts 3-, 4-, and 5-carbon skeletons, especially from the breakdown of amino acids, at other points in the pathway. For example, when deaminated, the amino acids aspartate and glutamate become the cycle intermediates oxaloacetate and α-ketoglutarate, respectively.
The Citric Acid Cycle Serves in Both Catabolic and Anabolic Processes
In aerobic organisms, the citric acid cycle is an amphibolic pathway, one that serves in both catabolic and anabolic processes. In the cycle’s anabolic role, oxaloacetate and α-ketoglutarate can be withdrawn from the cycle to serve as precursors of aspartate and glutamate by simple transamination (Chapter 22). Through aspartate and glutamate, the carbons of oxaloacetate and α-ketoglutarate are then used to build other amino acids, as well as purine and pyrimidine nucleotides. Succinyl-CoA is a central intermediate in the synthesis of the porphyrin ring of heme groups, which serve as oxygen carriers (in hemoglobin and myoglobin) and electron carriers (in cytochromes) (see Figs. 22-25, 22-26). Finally, oxaloacetate can be converted to glucose via gluconeogenesis (see Fig. 14-16).
One biosynthetic process is not possible for animals: the conversion of acetate or acetyl-CoA to glucose. Given that the carbon atoms of acetate molecules entering the citric acid cycle appear eight steps later in oxaloacetate, it might seem that the cycle could generate oxaloacetate from acetate, then the oxaloacetate could be used to synthesize glucose by gluconeogenesis. However, there is no net conversion of acetate to oxaloacetate; for every two carbons that enter the cycle as acetate (acetyl-CoA), two leave as . In bacteria, plants, fungi, and protists, another reaction sequence, the glyoxylate cycle, serves as a mechanism for converting acetate to carbohydrate. The glyoxylate cycle, which shares some reactions with the citric acid cycle, converts two molecules of acetate to one of oxaloacetate in a variant of the citric acid cycle in which the two decarboxylation steps are bypassed (see Fig. 20-45). Thus, plants and many simpler organisms can synthesize glucose from fatty acids, but humans and other animals cannot.
Under most circumstances, there is a dynamic steady state between reactions that siphon intermediates away from the citric acid cycle and those that supply additional carbon skeletons. When the withdrawal of cycle intermediates for use in biosynthesis lowers the concentrations of citric acid cycle intermediates enough to slow the cycle, the intermediates are replenished by anaplerotic reactions (Greek, “to refill”) (Fig. 16-15). Table 16-2 shows the most common anaplerotic reactions, all of which, in various tissues and organisms, convert either pyruvate or phosphoenolpyruvate to oxaloacetate or malate. The most important anaplerotic reaction in mammalian liver, kidney, and brown adipose tissue is the reversible carboxylation of pyruvate by to form oxaloacetate, catalyzed by pyruvate carboxylase. The enzymatic addition of a carboxyl group to pyruvate requires energy, which is supplied by ATP.
FIGURE 16-15 Role of the citric acid cycle in anabolism. Intermediates of the citric acid cycle are drawn off as precursors in many biosynthetic pathways. Shown in red are four anaplerotic reactions that replenish depleted cycle intermediates (see Table 16-2).
TABLE 16-2 Anaplerotic Reactions
Reaction
Tissue(s)/organism(s)
Liver, kidney
Heart, skeletal muscle
Higher plants, yeast, bacteria
Widely distributed in eukaryotes and bacteria
Pyruvate carboxylase is a regulatory enzyme and is virtually inactive in the absence of acetyl-CoA, its positive allosteric modulator. Whenever acetyl-CoA, the fuel for the citric acid cycle, is present in excess, it stimulates the pyruvate carboxylase reaction to produce more oxaloacetate, enabling the cycle to use more acetyl-CoA in the citrate synthase reaction.
Biotin in Pyruvate Carboxylase Carries One-Carbon () Groups
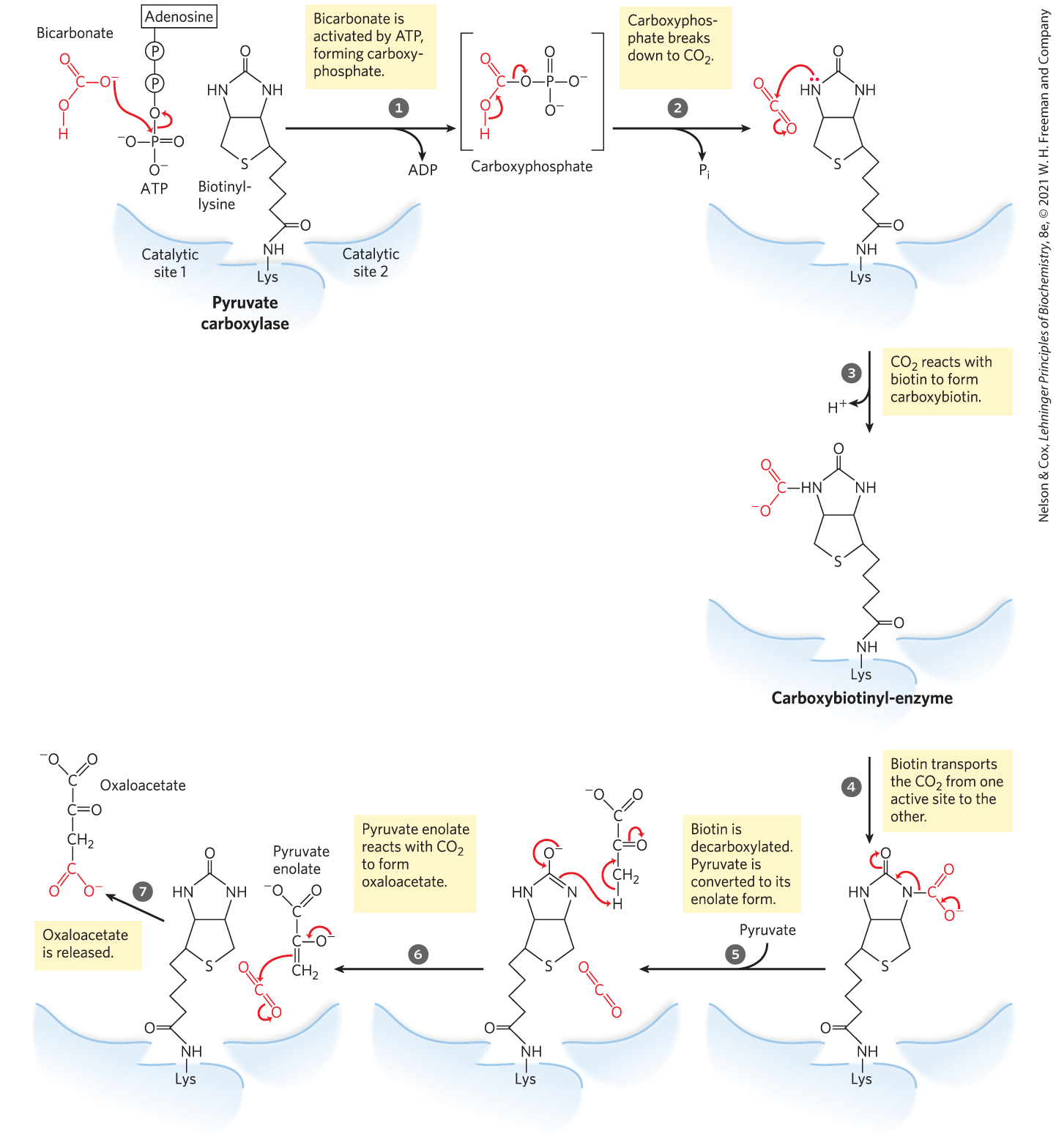
The pyruvate carboxylase reaction requires the vitamin biotin (Fig. 16-16), which is the prosthetic group of the enzyme. Biotin, which plays a key role in many carboxylation reactions, is a specialized carrier of one-carbon groups in their most oxidized form: . (The transfer of one-carbon groups in more reduced forms is mediated by other cofactors, notably tetrahydrofolate and S-adenosylmethionine, as described in Chapter 18.) Carboxyl groups are activated in a reaction that consumes ATP and joins to enzyme-bound biotin. This “activated” is then passed to an acceptor (pyruvate in this case) in a carboxylation reaction.
MECHANISM FIGURE 16-16 The role of biotin in the reaction catalyzed by pyruvate carboxylase. Biotin is attached to the enzyme through an amide bond with the ε-amino group of a Lys residue, forming biotinyl-enzyme. Biotin-mediated carboxylation reactions occur in two phases, generally catalyzed in separate active sites on the enzyme, as exemplified by the pyruvate carboxylase reaction. In the first phase (steps to ), bicarbonate is converted to the more activated , and then used to carboxylate biotin. The biotin acts as a carrier to transport the from one active site to another on an adjacent monomer of the tetrameric enzyme (step ). In the second phase (steps to ), catalyzed in this second active site, the reacts with pyruvate to form oxaloacetate.
Pyruvate carboxylase has four identical subunits, each containing a molecule of biotin covalently attached through an amide linkage to the ε-amino group of a specific Lys residue in the enzyme active site. Carboxylation of pyruvate proceeds in two steps (Fig. 16-16): first, a carboxyl group derived from is attached to biotin, then the carboxyl group is transferred to pyruvate to form oxaloacetate. These two steps occur at separate active sites; the long flexible arm of biotin transfers activated carboxyl groups from the first active site (on one monomer of the tetramer) to the second (on the adjacent monomer), functioning much like the long lipoyllysyl arm of in the PDH complex (Fig. 16-6) and the long arm of the CoA-like moiety in the acyl carrier protein involved in fatty acid synthesis (see Fig. 21-5); these are compared in Figure 16-17. Lipoate, biotin, and pantothenate all enter cells on the same transporter; all become covalently attached to proteins by similar reactions; and all provide a flexible tether that allows bound reaction intermediates to move from one active site to another in an enzyme complex, without dissociating from it. That is, all participate in substrate channeling.
FIGURE 16-17 Biological tethers. The cofactors lipoate, biotin, and the combination of β-mercaptoethylamine and pantothenate form long, flexible arms (green) on the enzymes to which they are covalently bound, acting as tethers that move intermediates from one active site to the next. The group shaded light red is, in each case, the point of attachment of the activated intermediate to the tether.
SUMMARY 16.3 The Hub of Intermediary Metabolism
The citric acid cycle is amphibolic, serving in both catabolism and anabolism. Besides acetyl-CoA, any compound that gives rise to a four- or five-carbon intermediate of the citric acid cycle — for example, the breakdown products of many amino acids — can be oxidized by the cycle. Cycle intermediates can be drawn off and used as the starting material for a variety of biosynthetic products.
When intermediates are shunted from the citric acid cycle to other pathways, they are replenished by several anaplerotic reactions, which produce four-carbon intermediates by carboxylation of three-carbon compounds; pyruvate carboxylase is a major anaplerotic enzyme.
Enzymes that catalyze carboxylations commonly employ biotin to activate and to carry it to acceptors such as pyruvate or phosphoenolpyruvate.
 The cycle also accepts 3-, 4-, and 5-carbon skeletons, especially from the breakdown of amino acids, at other points in the pathway. For example, when deaminated, the amino acids aspartate and glutamate become the cycle intermediates oxaloacetate and α-ketoglutarate, respectively.
The cycle also accepts 3-, 4-, and 5-carbon skeletons, especially from the breakdown of amino acids, at other points in the pathway. For example, when deaminated, the amino acids aspartate and glutamate become the cycle intermediates oxaloacetate and α-ketoglutarate, respectively.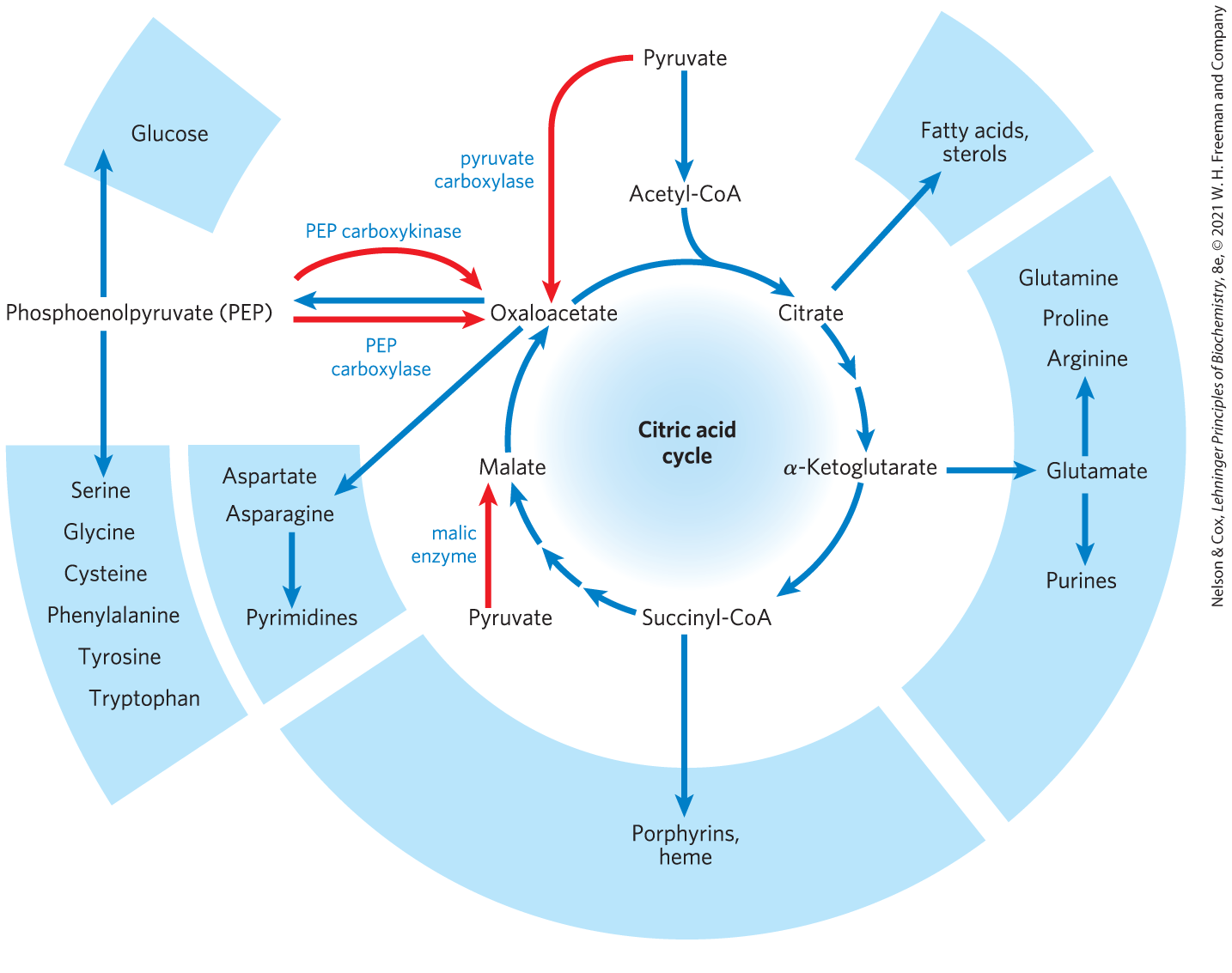

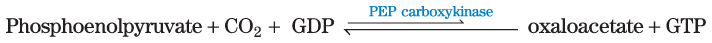

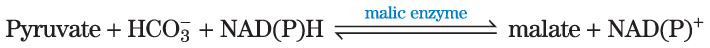
 to
to  ), bicarbonate is converted to the more activated
), bicarbonate is converted to the more activated  ). In the second phase (steps
). In the second phase (steps  to
to  ), catalyzed in this second active site, the reacts with pyruvate to form oxaloacetate.
), catalyzed in this second active site, the reacts with pyruvate to form oxaloacetate. These two steps occur at separate active sites; the long flexible arm of biotin transfers activated carboxyl groups from the first active site (on one monomer of the tetramer) to the second (on the adjacent monomer), functioning much like the long lipoyllysyl arm of in the PDH complex (
These two steps occur at separate active sites; the long flexible arm of biotin transfers activated carboxyl groups from the first active site (on one monomer of the tetramer) to the second (on the adjacent monomer), functioning much like the long lipoyllysyl arm of in the PDH complex (
 The citric acid cycle is amphibolic, serving in both catabolism and anabolism. Besides acetyl-CoA, any compound that gives rise to a four- or five-carbon intermediate of the citric acid cycle — for example, the breakdown products of many amino acids — can be oxidized by the cycle. Cycle intermediates can be drawn off and used as the starting material for a variety of biosynthetic products.
The citric acid cycle is amphibolic, serving in both catabolism and anabolism. Besides acetyl-CoA, any compound that gives rise to a four- or five-carbon intermediate of the citric acid cycle — for example, the breakdown products of many amino acids — can be oxidized by the cycle. Cycle intermediates can be drawn off and used as the starting material for a variety of biosynthetic products.