As we saw in Chapter 14 (Fig. 14-9), glycogen obtained in the diet is broken down by α-amylases, hydrolytic enzymes that act in the mouth and gut to convert glycogen to free glucose. (Dietary starch is hydrolyzed in a similar way.) But glycogen stored in cells (endogenous glycogen) is degraded by a different pathway. We begin with the breakdown of cellular glycogen to glucose 1-phosphate (glycogenolysis), then turn to synthesis of glycogen (glycogenesis).
Glycogen Breakdown Is Catalyzed by Glycogen Phosphorylase
In skeletal muscle and liver, the glucose units of the outer branches of glycogen enter the glycolytic pathway through the action of three enzymes: glycogen phosphorylase, glycogen debranching enzyme, and phosphoglucomutase. Glycogen phosphorylase catalyzes the reaction in which an glycosidic linkage between two glucose residues at a nonreducing end of glycogen undergoes attack by inorganic phosphate , removing the terminal glucose residue as α-d-glucose 1-phosphate (Fig. 15-3). This phosphorolysis reaction is different from the hydrolysis of glycosidic bonds by amylase during intestinal degradation of dietary glycogen and starch. In phosphorolysis, some of the energy of the glycosidic bond is conserved in the formation of the phosphate ester glucose 1-phosphate.
FIGURE 15-3 Removal of a glucose residue from the nonreducing end of a glycogen chain by glycogen phosphorylase. This process is repetitive; the enzyme removes successive glucose residues, creating a new nonreducing end, until it reaches the fourth glucose unit from a branch point (see Fig. 15-4).
Pyridoxal phosphate is an essential cofactor in the glycogen phosphorylase reaction. It is covalently attached near the enzyme active site, where its phosphate group acts as a general acid catalyst, promoting attack by on the glycosidic bond. (This is an unusual role for pyridoxal phosphate; its more typical role is as a cofactor in amino acid metabolism; see Fig. 18-6.)
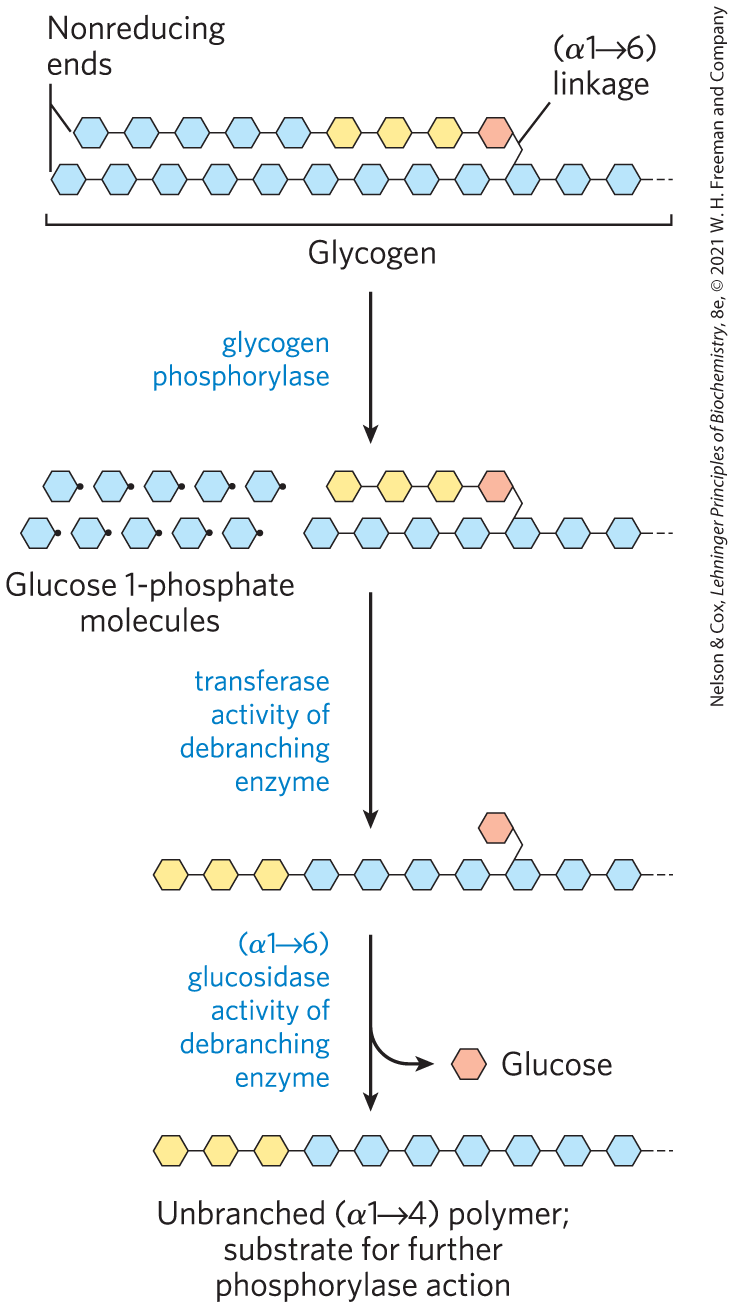
Glycogen phosphorylase acts repetitively on the nonreducing ends of glycogen branches until it reaches a point four glucose residues away from an branch point, where its action stops. Further degradation by glycogen phosphorylase can occur only after the debranching enzyme, formally known as oligo to glucantransferase, catalyzes two successive reactions that transfer branches (Fig. 15-4) to form straight chains. Once these branches are transferred and the glucosyl residue at C-6 is hydrolyzed, glycogen phosphorylase activity can continue.
FIGURE 15-4 Glycogen breakdown near an branch point. Following sequential removal of terminal glucose residues by glycogen phosphorylase (see Fig. 15-3), glucose residues near a branch are removed in a two-step process that requires a bifunctional debranching enzyme. First, the transferase activity of the enzyme shifts a block of three glucose residues from the branch to a nearby nonreducing end, to which the segment is reattached in linkage. The single glucose residue remaining at the branch point, in linkage, is then released as free glucose by the glucosidase activity of the debranching enzyme. The glucose residues are shown in shorthand form.
Glucose 1-Phosphate Can Enter Glycolysis or, in Liver, Replenish Blood Glucose
Glucose 1-phosphate, the end product of the glycogen phosphorylase reaction, is converted to glucose 6-phosphate by phosphoglucomutase, which catalyzes the reversible reaction:
Initially phosphorylated at a Ser residue, the enzyme donates its phosphoryl group to C-6 of the substrate, then accepts a phosphoryl group from C-1 (Fig. 15-5).
FIGURE 15-5 Reaction catalyzed by phosphoglucomutase. The reaction begins with the enzyme phosphorylated on a Ser residue. In step , the enzyme donates its phosphoryl group (blue) to glucose 1-phosphate, producing glucose 1,6-bisphosphate. In step , the phosphoryl group at C-1 of glucose 1,6-bisphosphate (red) is transferred back to the enzyme, re-forming the phosphoenzyme and producing glucose 6-phosphate.
The glucose 6-phosphate formed from glycogen in skeletal muscle can enter glycolysis and serve as an energy source to support muscle contraction. In liver, glycogen breakdown serves a different purpose: to release glucose into the blood when the blood glucose level drops, as it does between meals. This requires the enzyme glucose 6-phosphatase, present in liver and kidney but not in other tissues. The enzyme is an integral protein of the endoplasmic reticulum, with its active site on the lumenal side of the ER. Glucose 6-phosphate formed in the cytosol is transported into the ER lumen by a specific transporter (T1) (Fig. 15-6) and hydrolyzed at the lumenal surface by glucose 6-phosphatase. The resulting and glucose are carried back into the cytosol by two different transporters (T2 and T3), and the glucose enters the blood via the plasma membrane transporter, GLUT2. Notice that by having the active site of glucose 6-phosphatase in the ER lumen, the cell separates this reaction from the process of glycolysis, which takes place in the cytosol and would be aborted by the action of glucose 6-phosphatase. Genetic defects in either glucose 6-phosphatase or T1 lead to serious derangement of glycogen metabolism, resulting in type Ia glycogen storage disease (Box 15-1).
FIGURE 15-6 Hydrolysis of glucose 6-phosphate by glucose 6-phosphatase of the liver ER. The catalytic site of glucose 6-phosphatase faces the lumen of the ER. A glucose 6-phosphate (G6P) transporter (T1) carries the substrate from the cytosol to the lumen, where glucose 6-phosphatase releases . The products, glucose and , pass to the cytosol on specific transporters (T2 and T3). Glucose leaves the cell via the GLUT2 transporter in the plasma membrane.
Because muscle and adipose tissue lack glucose 6-phosphatase, they cannot convert the glucose 6-phosphate formed by glycogen breakdown to glucose, and these tissues therefore do not contribute glucose to the blood.
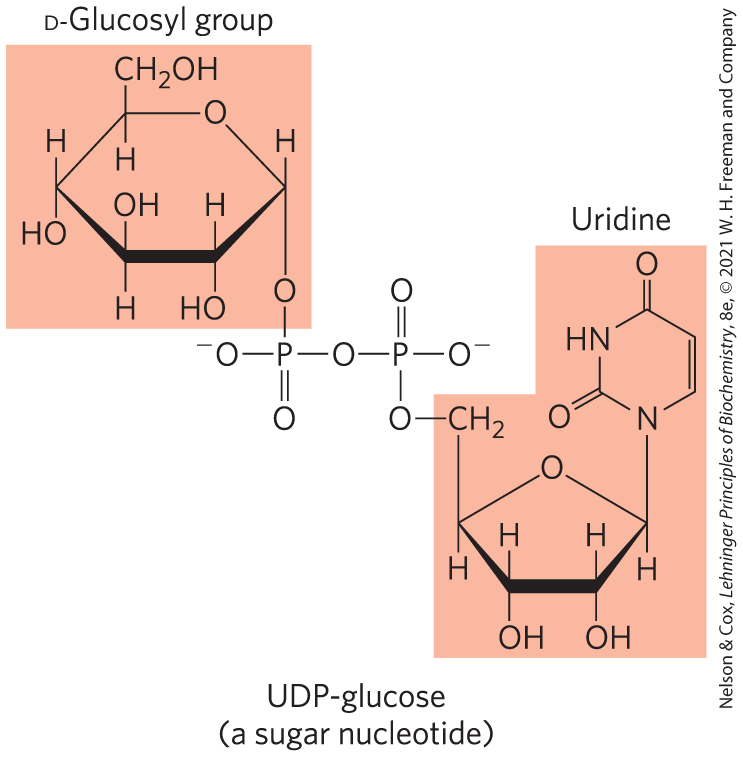
The Sugar Nucleotide UDP-Glucose Donates Glucose for Glycogen Synthesis
Many of the reactions in which hexoses are transformed or polymerized involve sugar nucleotides, compounds in which the anomeric carbon of a sugar is activated by attachment to a nucleotide through a phosphate ester linkage. Sugar nucleotides are the substrates for polymerization of monosaccharides into disaccharides, glycogen, starch, cellulose, and more complex extracellular polysaccharides. They are also key intermediates in the production of the aminohexoses and deoxyhexoses found in some of these polysaccharides, and in the synthesis of vitamin C (l-ascorbic acid). The role of sugar nucleotides in the biosynthesis of glycogen and many other carbohydrate derivatives was discovered in 1953 by the Argentine biochemist Luis Leloir.
The suitability of sugar nucleotides for biosynthetic reactions stems from several properties:
Their formation is metabolically irreversible, contributing to the irreversibility of the synthetic pathways in which they are intermediates. The condensation of a nucleoside triphosphate with a hexose 1-phosphate to form a sugar nucleotide has a small positive free-energy change, but the reaction releases , which is immediately hydrolyzed by inorganic pyrophosphatase (Fig. 15-7), in a reaction that is strongly exergonic . This keeps the cellular concentration of low, ensuring that the actual free-energy change in the cell is favorable. In effect, rapid removal of the product, driven by the large, negative free-energy change of hydrolysis, pulls the synthetic reaction forward. This is a common strategy in biological polymerization reactions.
Although the chemical transformations of sugar nucleotides do not involve the atoms of the nucleotide itself, the nucleotide moiety has many groups that can undergo noncovalent interactions with enzymes; the additional free energy of binding can contribute significantly to catalytic activity (Chapter 6; see also p. 294).
Like phosphate, the nucleotidyl group (UDP or ADP, for example) is an excellent leaving group, facilitating nucleophilic attack by activating the sugar carbon to which it is attached.
By “tagging” some hexoses with nucleotidyl groups, cells can set them aside in a pool for a particular purpose (glycogen synthesis, for example), separate from hexose phosphates destined for another purpose (such as glycolysis).
FIGURE 15-7 Formation of a sugar nucleotide. A condensation reaction occurs between a nucleoside triphosphate (NTP) and a sugar phosphate. The negatively charged oxygen on the sugar phosphate serves as a nucleophile, attacking the α phosphate of the nucleoside triphosphate and displacing pyrophosphate. The reaction is pulled in the forward direction by the hydrolysis of by inorganic pyrophosphatase.
Glycogen synthesis takes place in virtually all animal tissues but is especially prominent in the liver and skeletal muscles. The starting point for synthesis of glycogen is glucose 6-phosphate. This can be derived from free glucose in a reaction catalyzed by the isozymes hexokinase I and II in muscle and hexokinase IV (glucokinase) in liver:
However, some ingested glucose takes a more roundabout path to glycogen. It is first taken up by erythrocytes and converted to lactate via glycolysis; the lactate is then taken up by the liver and converted to glucose 6-phosphate by gluconeogenesis.
To start glycogen synthesis, the glucose 6-phosphate is converted to glucose 1-phosphate in the phosphogluco-mutase reaction:
The product is then converted to UDP-glucose by the action of UDP-glucose pyrophosphorylase, in a key step of glycogen biosynthesis:
Notice that this enzyme is named for the reverse reaction; in the cell, the reaction proceeds in the direction of UDP-glucose formation, because the pyrophosphate concentration is kept low by its immediate hydrolysis by inorganic pyrophosphatase (Fig. 15-7).
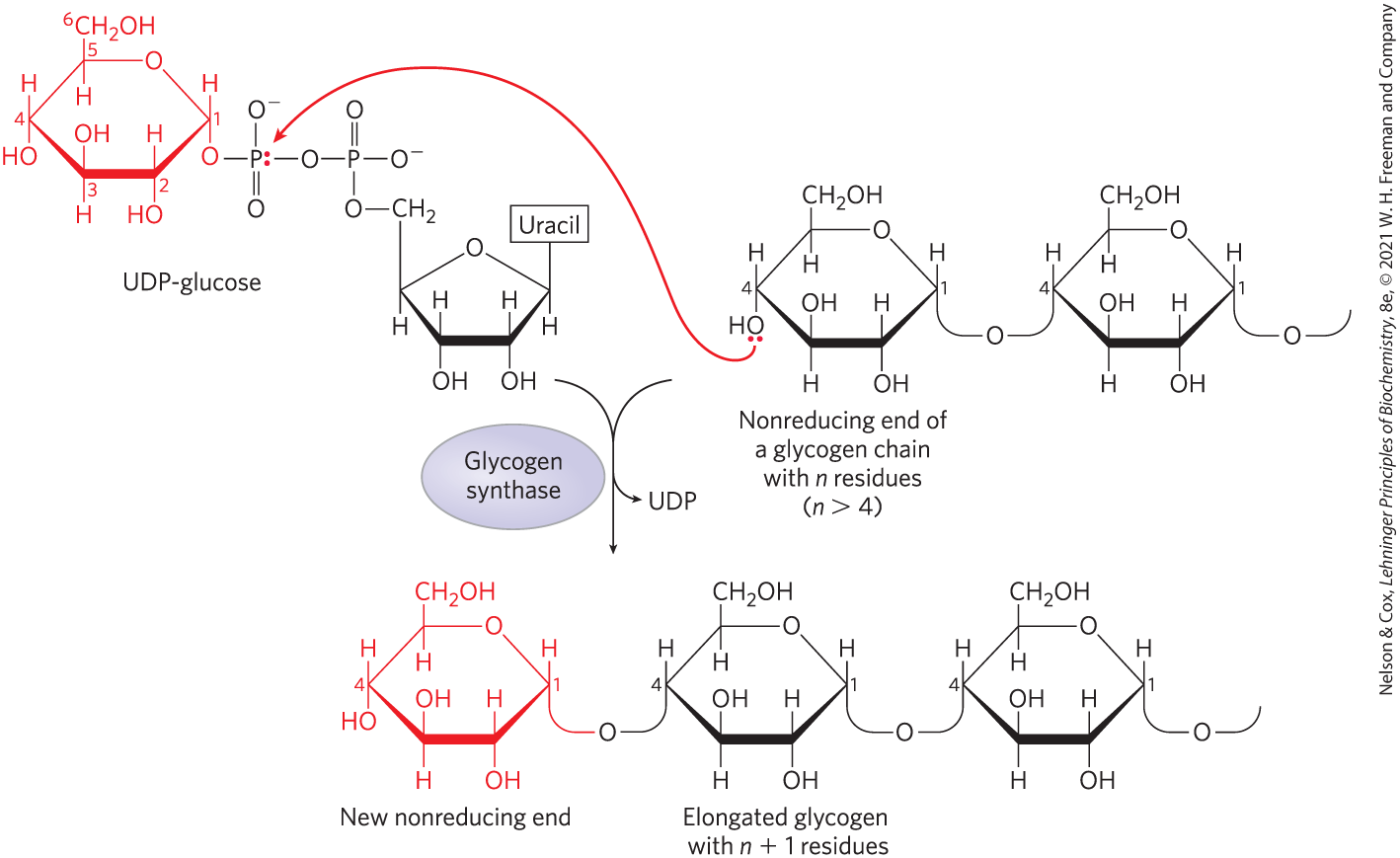
UDP-glucose is the immediate donor of glucose residues in the reaction catalyzed by glycogen synthase, which promotes the transfer of the glucose residue from UDP-glucose to a nonreducing end of a branched glycogen molecule, forming an linkage (Fig. 15-8). The overall equilibrium of the path from glucose 6-phosphate to glycogen lengthened by one glucose unit greatly favors synthesis of glycogen.
FIGURE 15-8 Glycogen synthesis. A glycogen chain is elongated by glycogen synthase. The enzyme transfers the glucose residue of UDP-glucose to the nonreducing end of a glycogen branch to make a new linkage.
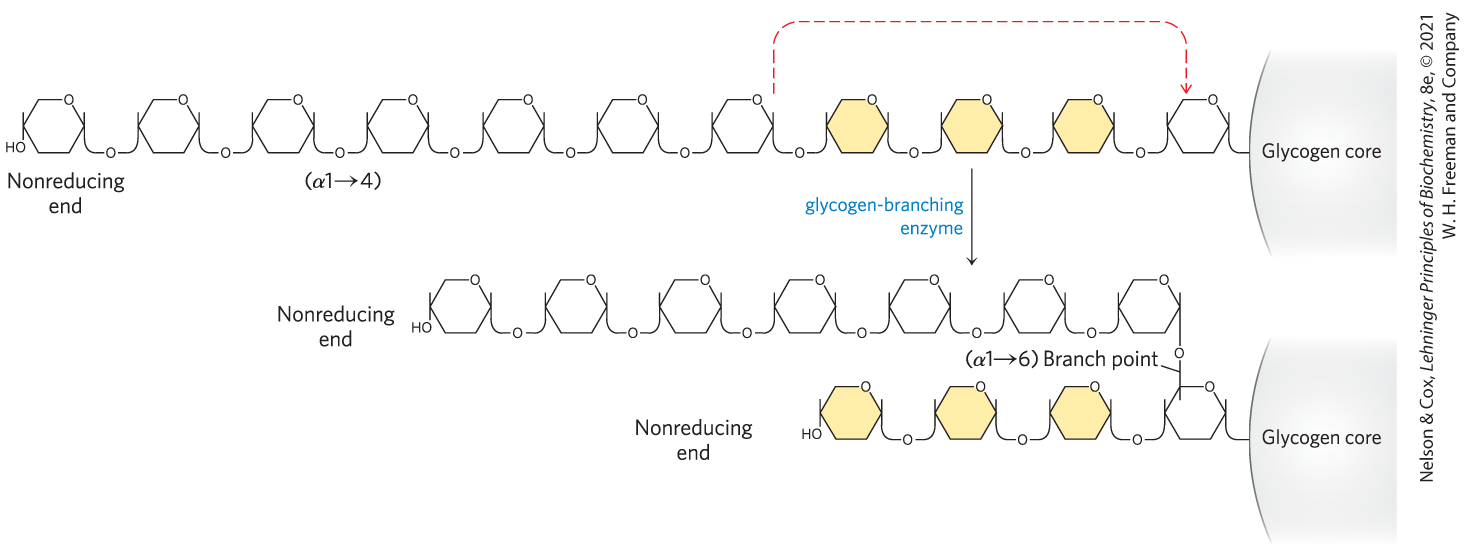
Glycogen synthase cannot make the bonds found at the branch points of glycogen; these are formed by the glycogen-branching enzyme, also called amylo to transglycosylase, or glycosyl transferase. The glycogen-branching enzyme catalyzes transfer of a terminal fragment of 6 or 7 glucose residues from the nonreducing end of a glycogen branch having at least 11 residues to the C-6 hydroxyl group of a glucose residue at a more interior position of the same or another glycogen chain, thus creating a new branch (Fig. 15-9). Further glucose residues may be added to the new branch by glycogen synthase. The biological effect of branching is to increase the number of nonreducing ends. This increases the number of sites accessible to glycogen phosphorylase and glycogen synthase, both of which act only at nonreducing ends.
FIGURE 15-9 Branch synthesis in glycogen. The glycogen-branching enzyme forms a new branch point during glycogen synthesis.
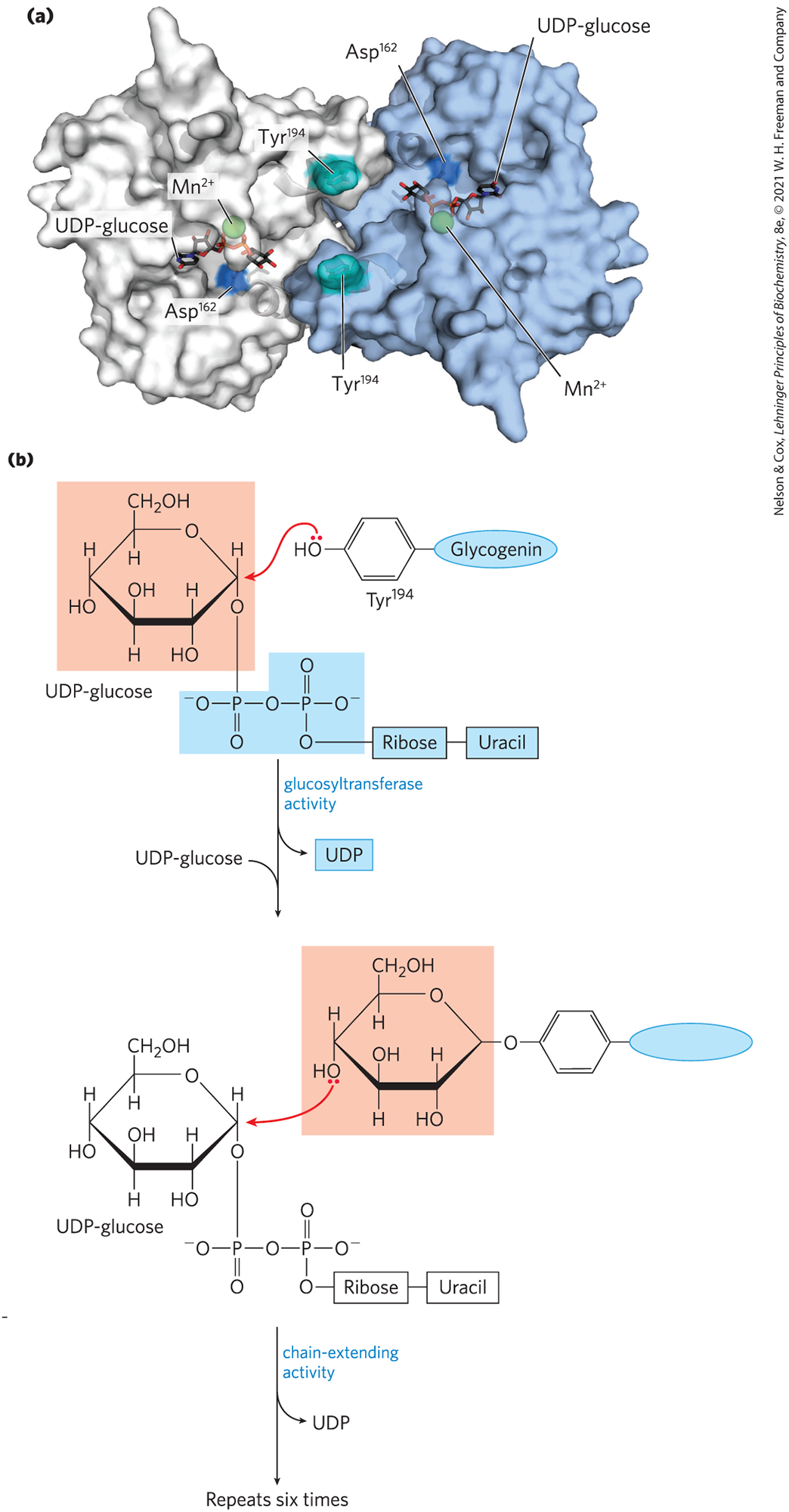
Glycogenin Primes the Initial Sugar Residues in Glycogen
Glycogen synthase cannot initiate a new glycogen chain de novo. It requires a primer, usually a preformed polyglucose chain. So, how is a new glycogen molecule initiated? The intriguing protein glycogenin (Fig. 15-10) is both the primer on which new chains are assembled and the enzyme that catalyzes their assembly. The first step in the synthesis of a new glycogen molecule is the transfer of a glucose residue from UDP-glucose to the hydroxyl group of of glycogenin. Each subunit of the glycogenin homodimer glycosylates of the other subunit. The glycosidic bond in the product has the same configuration about the C-1 of glucose as the substrate UDP-glucose, suggesting that the transfer of glucose from UDP to occurs in two steps. The first step is probably a nucleophilic attack by , forming a temporary intermediate with inverted configuration. A second nucleophilic attack by then restores the starting configuration. The nascent chains are extended by the sequential addition of seven more glucose residues, each derived from UDP-glucose; the reactions are catalyzed by the chain-extending activity of glycogenin. At this point, glycogen synthase takes over, further extending the glycogen chain. Glycogenin remains buried within the β-particle, covalently attached to the two reducing ends of the glycogen molecule.
FIGURE 15-10 Glycogenin. (a) The protein is a homodimer. The substrate, UDP-glucose, is bound in a region near the amino terminus and is some distance from the residues — 15 Å from the Tyr in the same monomer, 12 Å from the Tyr in the dimeric partner. Each UDP-glucose is bound through its phosphates to a ion, which is essential to catalysis. is believed to function as an electron-pair acceptor (Lewis acid) to stabilize the leaving group, UDP. (b) Glycogenin catalyzes two distinct reactions. Initial attack by the hydroxyl group of on C-1 of the glucosyl moiety of UDP-glucose results in a glucosylated Tyr residue. The C-1 of another UDP-glucose molecule is now attacked by the C-4 hydroxyl group of the terminal glucose, and this sequence repeats to form a nascent glycogen molecule of eight glucose residues attached by glycosidic linkages. [(a) Data from PDB ID 1LL2, B. J. Gibbons et al., J. Mol. Biol. 319:463, 2002.]
SUMMARY 15.2 Breakdown and Synthesis of Glycogen
Glycogen phosphorylase catalyzes phosphorolytic cleavage at the nonreducing ends of glycogen chains, producing glucose 1-phosphate. The debranching enzyme transfers branches onto main chains and releases the residue at the branch as free glucose.
Phosphoglucomutase interconverts glucose 1-phosphate and glucose 6-phosphate. Glucose 6-phosphate can enter glycolysis or, in liver, it can be converted to free glucose by glucose 6-phosphatase in the endoplasmic reticulum, then exported to replenish blood glucose.
The sugar nucleotide UDP-glucose donates glucose residues to the nonreducing end of glycogen in the reaction catalyzed by glycogen synthase, producing short -linked segments. A separate branching enzyme produces the linkages at branch points.
New glycogen particles begin with the autocatalytic formation of a glycosidic bond between the glucose of UDP-glucose and a Tyr residue of the protein glycogenin, followed by addition of several glucose residues to form a primer that can be acted on by glycogen synthase.
 In skeletal muscle and liver, the glucose units of the outer branches of glycogen enter the glycolytic pathway through the action of three enzymes: glycogen phosphorylase, glycogen debranching enzyme, and phosphoglucomutase. Glycogen phosphorylase catalyzes the reaction in which an glycosidic linkage between two glucose residues at a nonreducing end of glycogen undergoes attack by inorganic phosphate
In skeletal muscle and liver, the glucose units of the outer branches of glycogen enter the glycolytic pathway through the action of three enzymes: glycogen phosphorylase, glycogen debranching enzyme, and phosphoglucomutase. Glycogen phosphorylase catalyzes the reaction in which an glycosidic linkage between two glucose residues at a nonreducing end of glycogen undergoes attack by inorganic phosphate 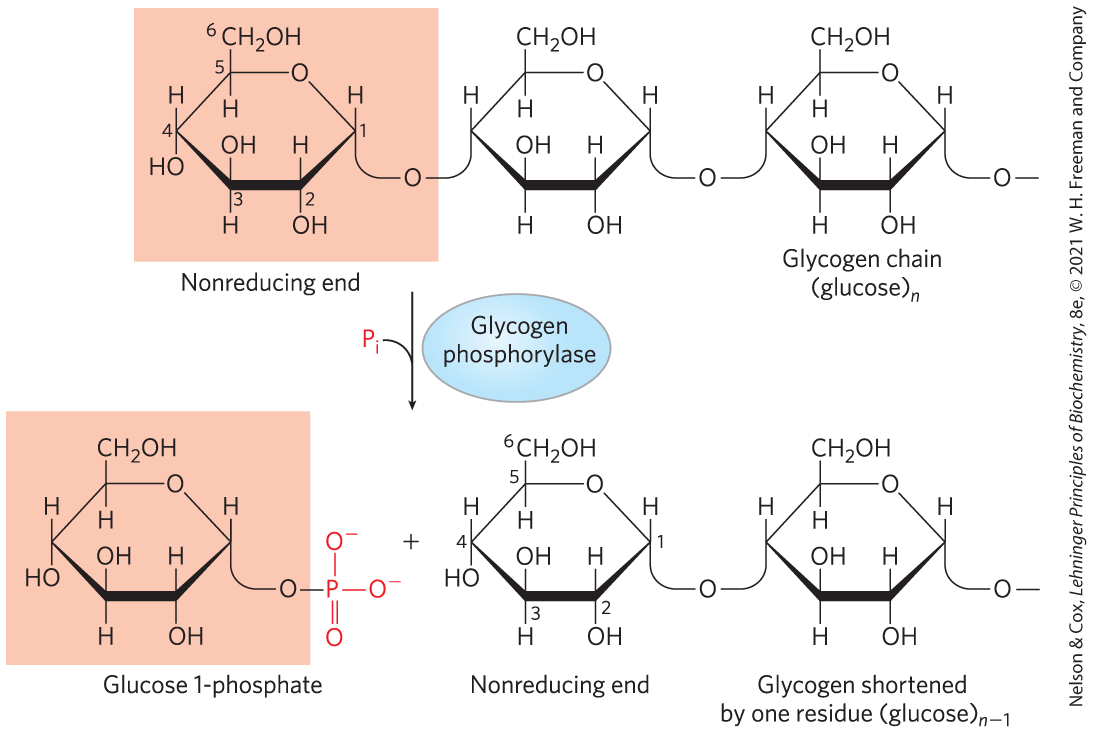
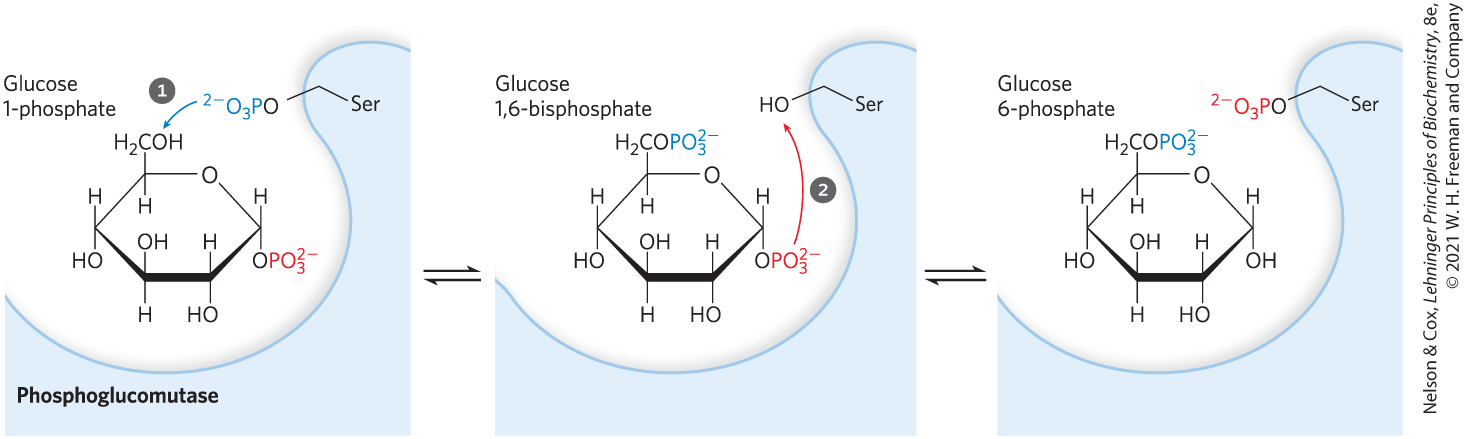
 , the enzyme donates its phosphoryl group (blue) to glucose 1-phosphate, producing glucose 1,6-bisphosphate. In step
, the enzyme donates its phosphoryl group (blue) to glucose 1-phosphate, producing glucose 1,6-bisphosphate. In step  , the phosphoryl group at C-1 of glucose 1,6-bisphosphate (red) is transferred back to the enzyme, re-forming the phosphoenzyme and producing glucose 6-phosphate.
, the phosphoryl group at C-1 of glucose 1,6-bisphosphate (red) is transferred back to the enzyme, re-forming the phosphoenzyme and producing glucose 6-phosphate.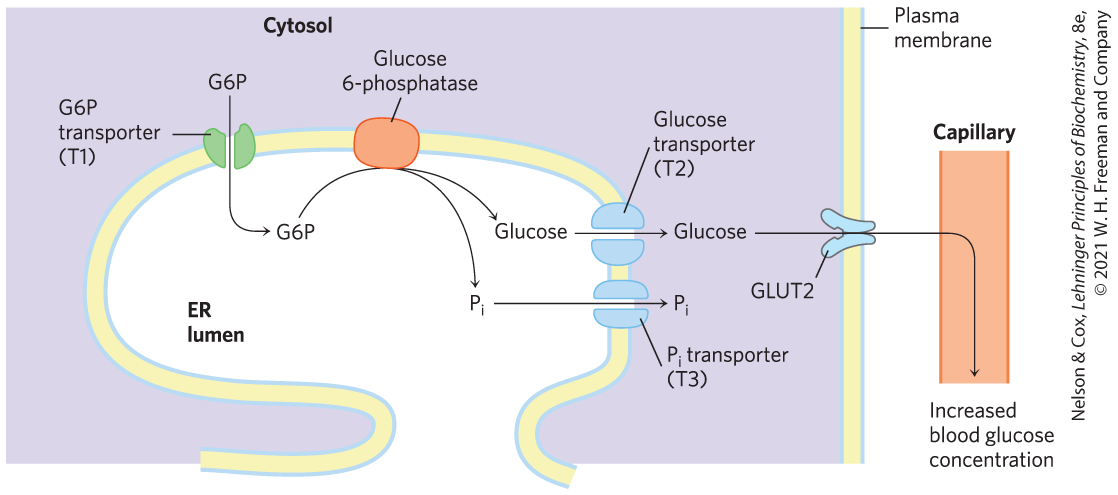
 The suitability of sugar nucleotides for biosynthetic reactions stems from several properties:
The suitability of sugar nucleotides for biosynthetic reactions stems from several properties: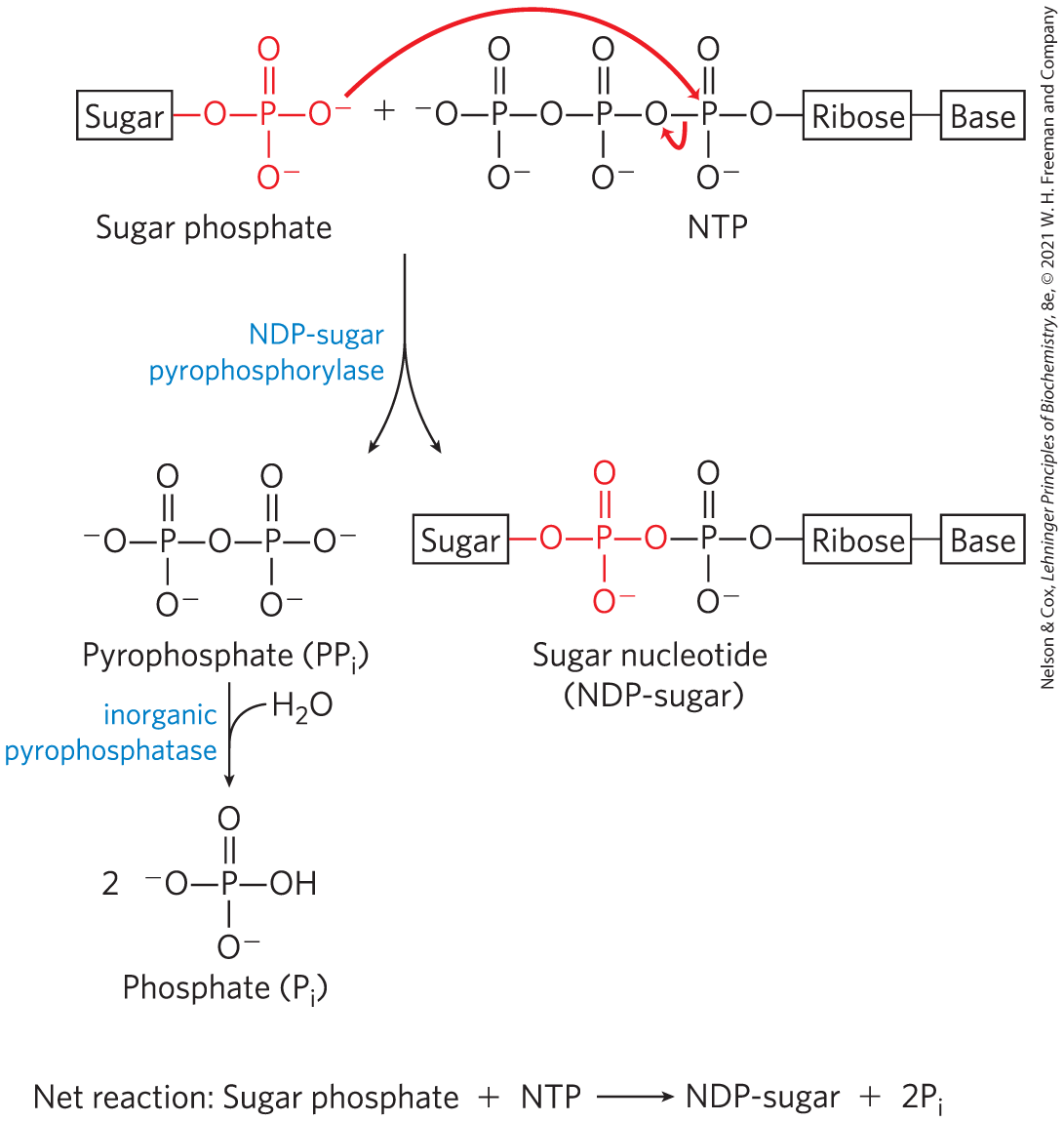
 Glycogen phosphorylase catalyzes phosphorolytic cleavage at the nonreducing ends of glycogen chains, producing glucose 1-phosphate. The debranching enzyme transfers branches onto main chains and releases the residue at the branch as free glucose.
Glycogen phosphorylase catalyzes phosphorolytic cleavage at the nonreducing ends of glycogen chains, producing glucose 1-phosphate. The debranching enzyme transfers branches onto main chains and releases the residue at the branch as free glucose.