Study Guide: Ion Channels & Membrane Electrophysiology
1. Overview
Ion flow is governed by:
Membrane permeability (via specific ion channels).
Concentration gradients (chemical driving force).
Electrical driving forces (voltage difference across the membrane).
Membrane potential (
Selective ion channels.
Active transport mechanisms (e.g., Na⁺/K⁺ pumps).
Impermeant anions and charge separation.
Equilibrium potential (Nernst) defines the voltage at which ion movement is balanced by the chemical gradient:
60 mV is an approximation at ~37 °C (more precisely 61–62 mV).
2. Key Equations
Ohm’s Law
Conductance (
Weighted Sum for Membrane Potential (multiple ions)
Ion Current
Net current depends on the difference (
General Flow Equation
Time Constant
Chord Conductance
3. Capacitance & RC Properties
Capacitor stores charge by separating it across an insulating layer.
The cell membrane behaves as a capacitor:
Intracellular/extracellular fluid = conductors.
Lipid bilayer = dielectric (insulator).
3.1 Stored Charge
3.2 Capacitive Current
Faster change in voltage
3.3 Time Constant (
Governs how fast the voltage changes in response to current:
Time to reach
4. Measuring & Calculating Membrane Properties
Measure the Time Constant (
Inject a small current step; record time to reach
Measure Steady-State Voltage Response
After current injection, at steady-state:
Calculate Capacitance
Determine Cell Area
Typical biological membranes have
Find Specific Membrane Resistance
5. Membrane as an Electrical Circuit
Capacitor: lipid bilayer.
Resistor: ion channels.
Battery: Nernst potential (ionic gradient).
5.1 Equivalent Circuit
Membrane = RC circuit:
Capacitance (
Ohm’s Law:
Input Resistance (
5.2 Time Constant
Time to charge/discharge the membrane by 63%.
5.3 Length Constant
For cable-like processes (axon, dendrite):
6. Voltage-Gated Channels
Structure:
S4 segment contains positively charged amino acids (voltage sensor).
Ion Selectivity:
Channels discriminate ions based on size, charge, hydration.
Voltage Dependence:
Voltage sensors respond to membrane potential changes → channel gates open/close.
6.1 Na⁺ Channels
Fast activation and fast inactivation.
Inactivation ensures transient current even with sustained depolarization.
Refractory Period:
After an action potential, channels remain inactivated briefly → cannot reopen immediately.
6.2 K⁺ Channels
Slower activation (classical Hodgkin-Huxley channels).
Often do not inactivate (or do so minimally) → can remain open with prolonged depolarization.
Inward Rectifiers: block outward flow at positive potentials → stabilize resting potential.
6.3 Ca²⁺ Channels
Depolarization opens Ca²⁺ channels:
Often slower inactivation → can prolong action potentials (e.g., cardiac cells).
Ca²⁺ entry can trigger diverse intracellular events (secretion, contraction, signaling).
7. Current–Voltage (I–V) Relationships
Plotting current (
Reversal potential (where net current = 0).
Slope conductance =
Chord Conductance (
I–V Plot Regions:
Positive slope: typical ohmic or outward K⁺ flow.
Negative slope: inward rectification or channel inactivation phenomena.
8. Conductance–Voltage (G–V) Plots
Derived from I–V data:
Sigmoidal activation curve:
Half-activation voltage (
9. Single-Channel Behavior
Microscopic currents: discrete channel openings/closings.
Macroscopic current: sum of many channels.
Stochastic gating: channels open with certain probability depending on voltage, time, etc.
10. Protocols to Study Na⁺ Activation/Inactivation
Activation:
Clamp the membrane at various depolarizing steps from a negative holding potential.
Measure how quickly and strongly Na⁺ current activates.
Inactivation:
Use prepulses at different voltages.
Then step to a test voltage → measure available Na⁺ current.
Plot fraction of channels available vs. prepulse voltage → midpoint of inactivation.
11. Consequences of Na⁺ Channel Inactivation
Transient Na⁺ current despite sustained depolarization.
Refractory period ensures:
Unidirectional action potential propagation.
Finite firing frequency (cannot immediately trigger another AP).
Prolonged depolarization → channels remain inactivated → no further firing (weakness or paralysis).
12. External Ion Concentrations
Decreasing
Increasing
13. Pathophysiology: Muscle Weakness & Inexcitability
Shifts in Resting
Abnormal channel gating or ion gradients → altered excitability.
Input Resistance & Time Constant Changes:
Affected by channel density or membrane properties → can require bigger/longer stimuli to reach threshold.
Clinical Correlates:
Periodic paralysis, channelopathies → result from subtle gating alterations or ionic imbalances.
14. Axonal Conduction
14.1 Passive Spread (Cable Properties)
Voltage decays over distance if no regenerative channels are present.
Larger diameter → lower axial resistance (
14.2 Unmyelinated Axons
Continuous conduction:
Slower (few m/s in small fibers).
Speed often scales with axon diameter (e.g., squid giant axon ~20 m/s).
14.3 Myelinated Axons
Saltatory conduction:
Na⁺ channels clustered at nodes of Ranvier.
Myelin → lowers capacitance, increases membrane resistance in internodes.
Conduction speeds can reach ~100 m/s in large myelinated fibers.
| Property | Unmyelinated Axons | Myelinated Axons |
|---|---|---|
| Conduction Type | Continuous | Saltatory |
| Speed | Slow (~0.5–2 m/s) | Fast (~50–100 m/s) |
| Na⁺ Channel Location | Distributed | Clustered at nodes |
| Capacitance | High | Low |
| Resistance | Low | High |
Myelin reduces capacitance → less charge is needed to depolarize the next node.
15. Calcium Action Potentials
Ca²⁺ Channels:
Open upon depolarization, often inactivate more slowly → longer-duration APs.
Replacing Na⁺ with Ca²⁺:
Yields slower, longer spikes due to lower channel conductance and slower inactivation.
Cardiac Action Potentials:
Plateau phase from sustained Ca²⁺ influx.
Increasing
K⁺ currents eventually repolarize the cell.
Summary
Membrane potential depends on ion gradients, selective permeability, and ion channel gating.
RC properties (resistance
Time constant (
Voltage-gated channels underlie action potentials and are finely regulated by voltage sensors.
Ion concentrations critically affect excitability and AP amplitude.
Myelination optimizes conduction velocity without drastically increasing axon diameter.
Inactivation of channels (especially Na⁺) is key to the refractory period and directional AP propagation.
Properties of Voltage-Activated Calcium Channels
Subunit Structure :
main
has 4 domains
the main pore-forming subunit that determines ion selectivity, conductance, and gating properties
Four-transmembrane-spanning protein, initially found in skeletal muscle calcium channels but also present in the brain (e.g., stargazin).
Cytoplasmic protein that binds to the α1 subunit’s intracellular I-II linker, helping with trafficking to the plasma membrane and modulation
Derived from a single gene and proteolytically cleaved into an extracellular α2 portion and a membrane-spanning δ portion, which stabilizes the channel at the plasma membrane.
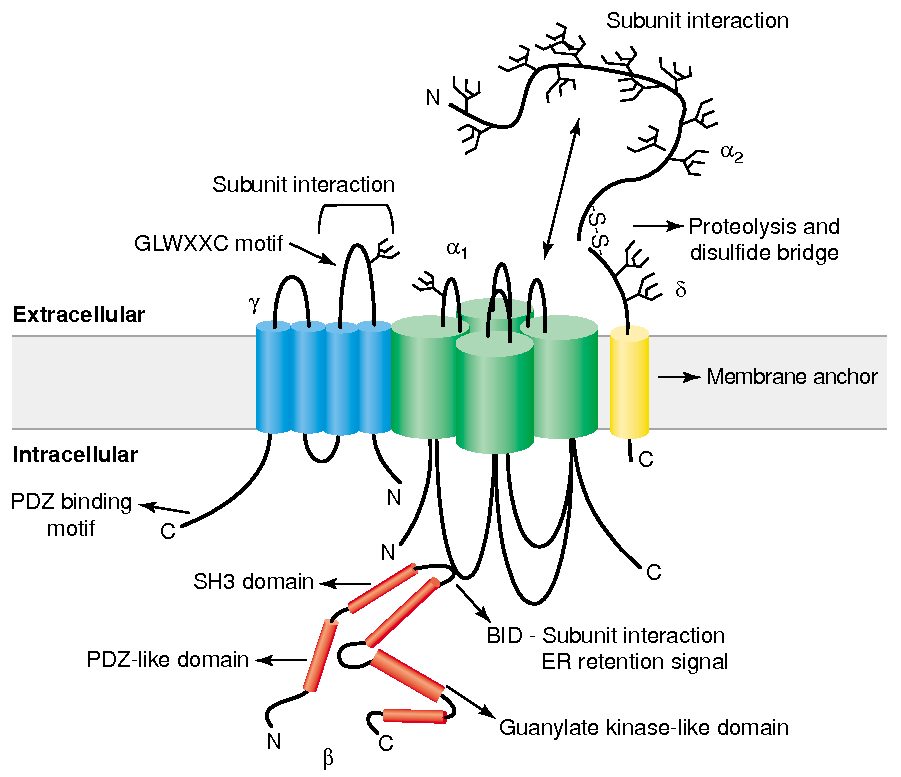
Gate / Voltage Sensor :
S4 segment :
Contains positively charged amino acids , arginine and lysine , every 3rd or 4th residue
unctions as the voltage sensor
Depolarization causes deformation in P-loops , creating a pore
Selectivity Filter for Calcium
Calcium selectivity is determined by the pore loop ( P-loop ) glutamates :
Glutamate residues ( E ) are conserved in each of the four domains ( I-IV ) of the α1 subunit
These residues form a high-affinity binding site for Ca2+ ,
ensuring preferential permeability over other ions
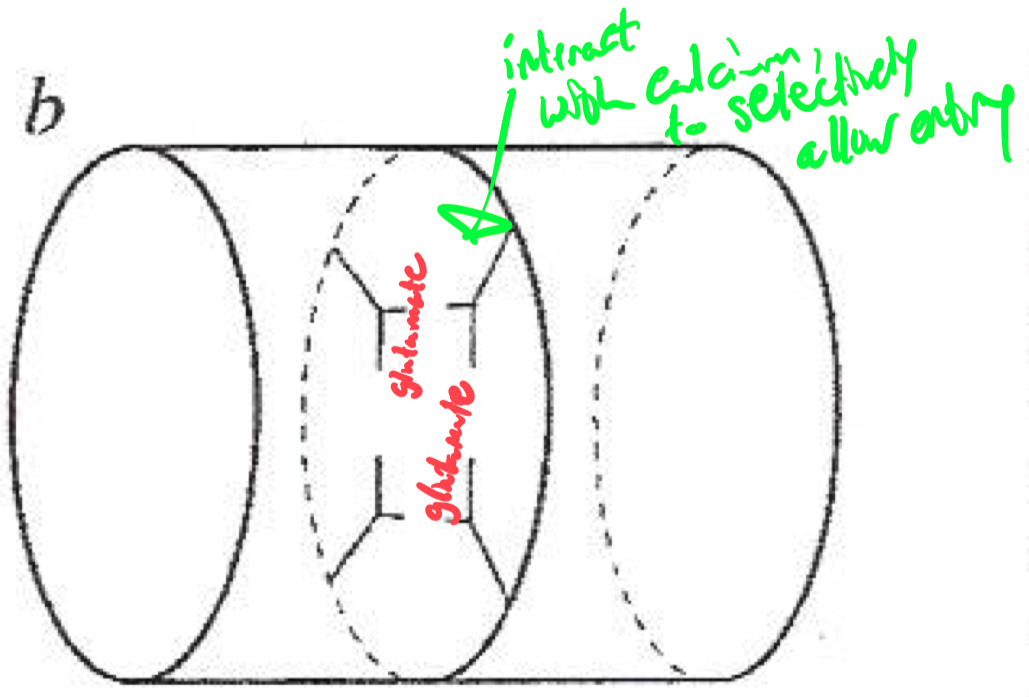
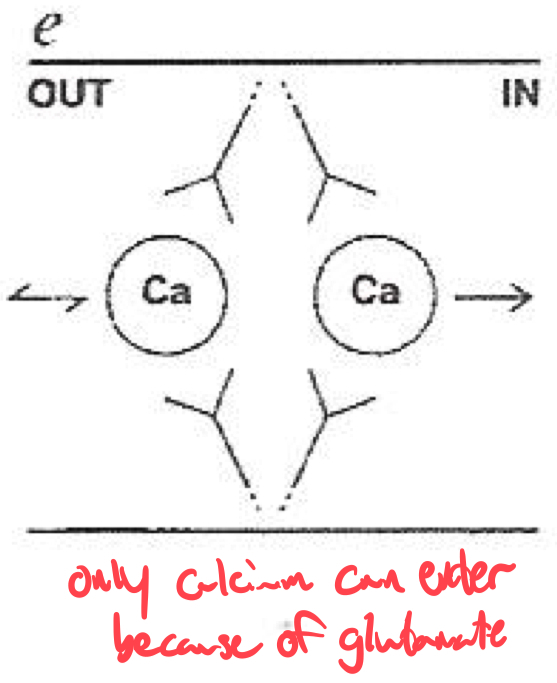
How would you classify or identify different subtypes of calcium channels ?
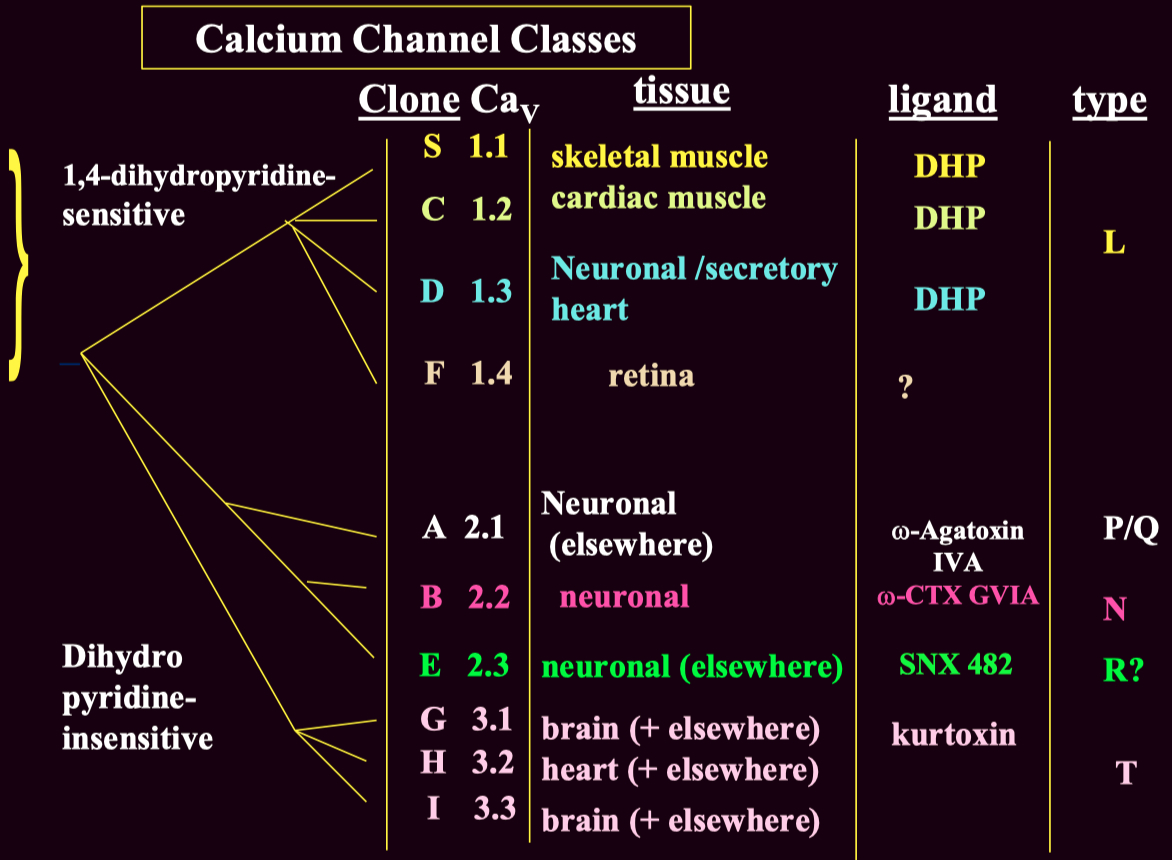
Voltage-gated calcium channels are classified into :
High-Voltage Activated ( HVA )
Low-Voltage Activated ( LVA ) ( T-type ) channels
These subtypes can be distinguished using :
Pharmacological blockers ( e.g., ω-Agatoxin IVA for P/Q-type channels )
Genetic expression ( different subtypes are present in different tissues )
Electrophysiology ( LVA T-type channels activate at lower voltages than HVA channels )
Channel Type Clone (CaV) Function Blockers L-type CaV1.1 - CaV1.4 Skeletal, cardiac muscle contraction, secretion DHP (nifedipine), verapamil P/Q-type CaV2.1 Neurotransmitter release (central) ω-Agatoxin IVA N-type CaV2.2 Neurotransmitter release (peripheral & central) ω-Conotoxin GVIA R-type CaV2.3 Uncertain SNX 482 T-type CaV3.1 - Cav3.3 Cell excitability Mibefradil, ethosuximide (controversial)
L-type blockers:
Dihydropyridines (DHPs): Nifedipine, Amlodipine.
Non-DHPs: Verapamil (cardiac selective).
P/Q-type blockers:
ω-Agatoxin IVA (funnel web spider toxin).
N-type blockers:
ω-Conotoxin GVIA (cone shell mollusk toxin).
Used for neuropathic pain management.
T-type blockers:
Mibefradil, ethosuximide (antiepileptics).
| Subunit | Type | Function |
|---|---|---|
| P/Q-type | CNS neurotransmitter release | |
| N-type | Neurotransmitter release (peripheral & central) | |
| L-type | Cardiac contraction | |
| L-type | Hormone secretion | |
| L-type | Skeletal muscle contraction | |
| T-type | Pacemaking, excitability |
Q: Explain why the resting
Q: Sketch/describe an RC charging curve and label
Q: On an
Q: How does changing
Q: Absolute vs. relative refractory periods in terms of Na
Q: Why might a muscle fiber at
Q: Raising
Q: A short current pulse doesn’t reach threshold, but a longer one does. Why? A: The membrane (RC circuit) needs enough time to charge up to threshold. A short pulse may end before hitting threshold.
Q: Multiple myelin wraps: effect on conduction velocity?
A: Myelin greatly reduces