In Chapter 10 we introduced two major classes of membrane phospholipids: glycerophospholipids and sphingolipids. Many different phospholipid species can be constructed by combining various fatty acids and polar head groups with the glycerol or sphingosine backbone (see Figs. 10-8, 10-11). All the biosynthetic pathways follow a few basic patterns. In general, the assembly of phospholipids from simple precursors requires (1) synthesis of the backbone molecule (glycerol or sphingosine); (2) attachment of fatty acid(s) to the backbone through an ester or amide linkage; (3) addition of a hydrophilic head group to the backbone through a phosphodiester linkage; and, in some cases, (4) alteration or exchange of the head group or the fatty acids to yield the final phospholipid product.
In eukaryotic cells, phospholipid synthesis occurs primarily on the surfaces of the smooth ER and the inner mitochondrial membrane. Some newly formed phospholipids remain at the site of synthesis, but most are destined for other cellular membranes. Once they arrive, phospholipids can be remodeled within membranes to alter the fatty acid constituents. The process by which water-insoluble phospholipids move from the site of synthesis to the point of their eventual function is not fully understood, but we discuss some mechanisms that have emerged in recent years.
Cells Have Two Strategies for Attaching Phospholipid Head Groups
Stage 1 of glycerophospholipid synthesis is shared with the pathway to triacylglycerols, the formation of glycerol 3-phosphate by one of the two paths shown in Fig. 21-17. In stage 2, fatty acyl groups are esterified to C-1 and C-2 of l-glycerol 3-phosphate to form phosphatidic acid. Usually, the fatty acid at C-1 is saturated and the one at C-2 is unsaturated. A second route to phosphatidic acid is the phosphorylation of a diacylglycerol by a specific kinase.
Eugene P. Kennedy, 1919–2011
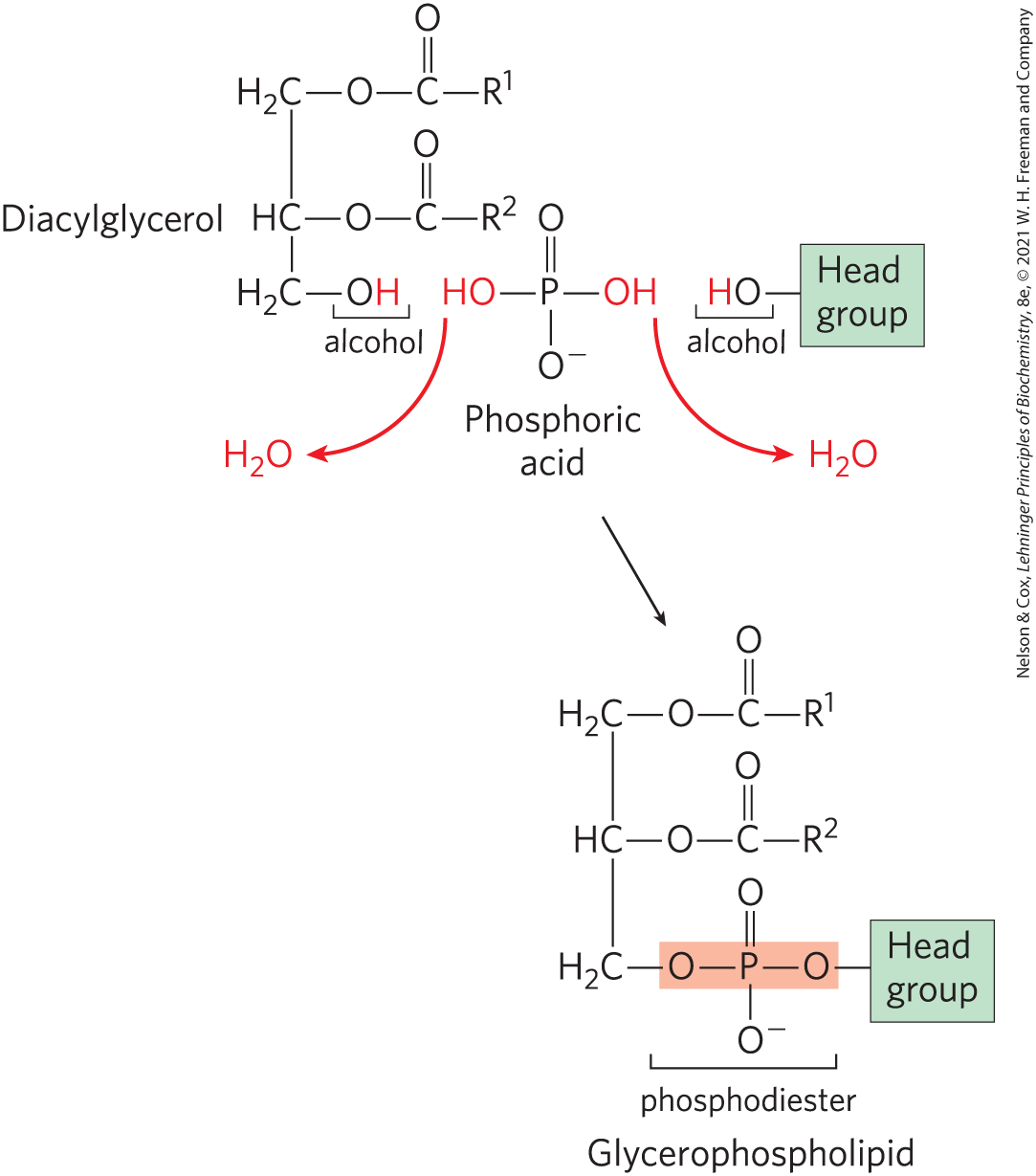
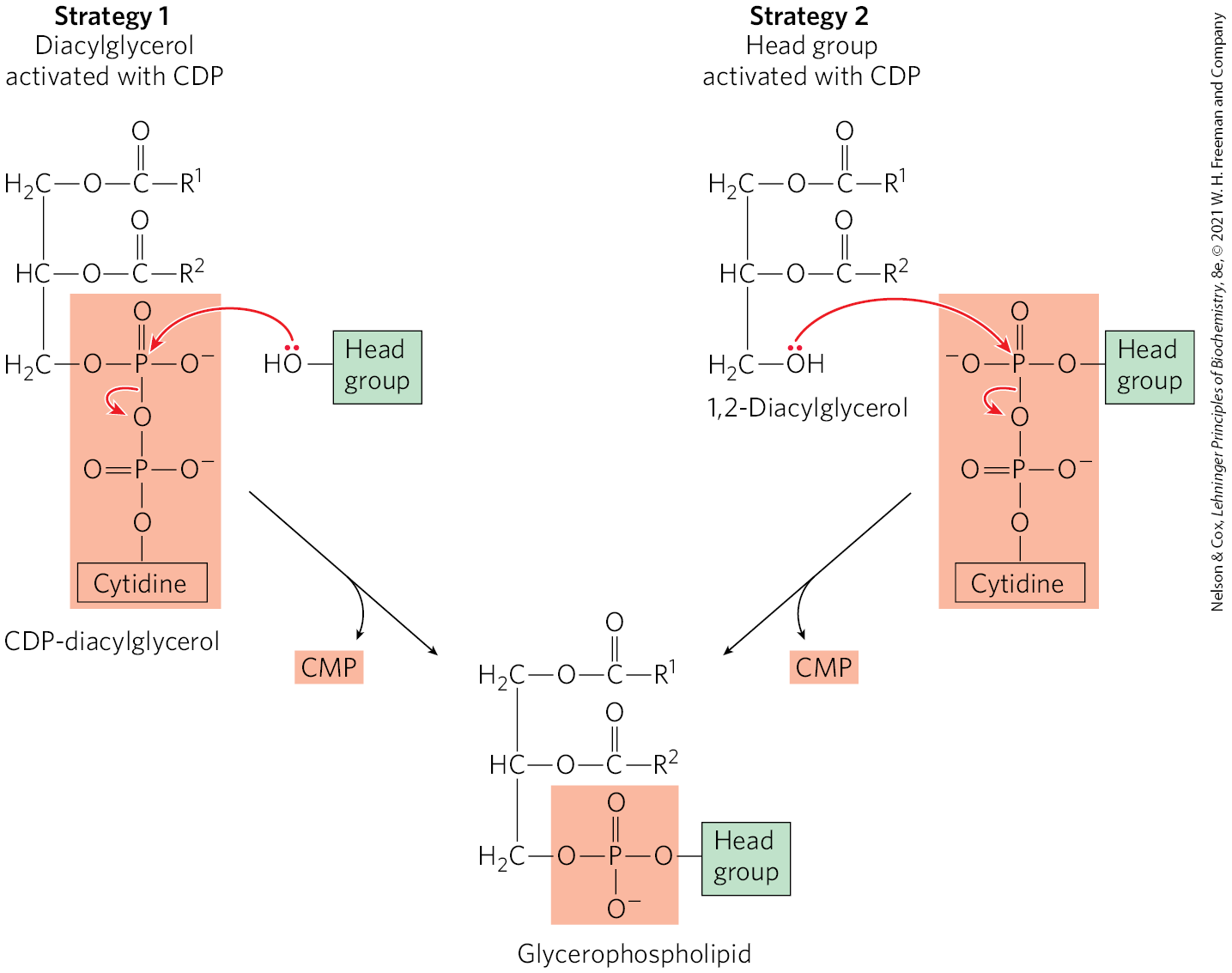
In stages 3 and 4, the polar head group of glycerophospholipids is attached through a phosphodiester bond, in which each of two alcohol hydroxyls (one on the polar head group and one on C-3 of glycerol) forms an ester with phosphoric acid (Fig. 21-23). In the biosynthetic process, one of the hydroxyls is first activated by attachment of a nucleotide, cytidine diphosphate (CDP). Cytidine monophosphate is then displaced in a nucleophilic attack by the other hydroxyl (Fig. 21-24). Two strategies are employed by mammals. The CDP is attached either to the diacylglycerol, forming the activated phosphatidic acid CDP-diacylglycerol (strategy 1), or to the hydroxyl of the head group (strategy 2). The central importance of cytidine nucleotides in lipid biosynthesis was discovered by Eugene P. Kennedy in the late 1950s, and this pathway is commonly referred to as the Kennedy pathway. In bacteria, only strategy 1 is used to generate glycerophospholipids.
FIGURE 21-23 Final stages of glycerophospholipid biosynthesis: head-group attachment. The phospholipid head group is attached to a diacylglycerol by a phosphodiester bond (shaded light red), formed when phosphoric acid condenses with two alcohols, eliminating two molecules of .
FIGURE 21-24 Two general strategies for forming the phosphodiester bond of phospholipids. In both cases, CDP supplies the phosphate group of the phosphodiester bond.
Pathways for Phospholipid Biosynthesis Are Interrelated
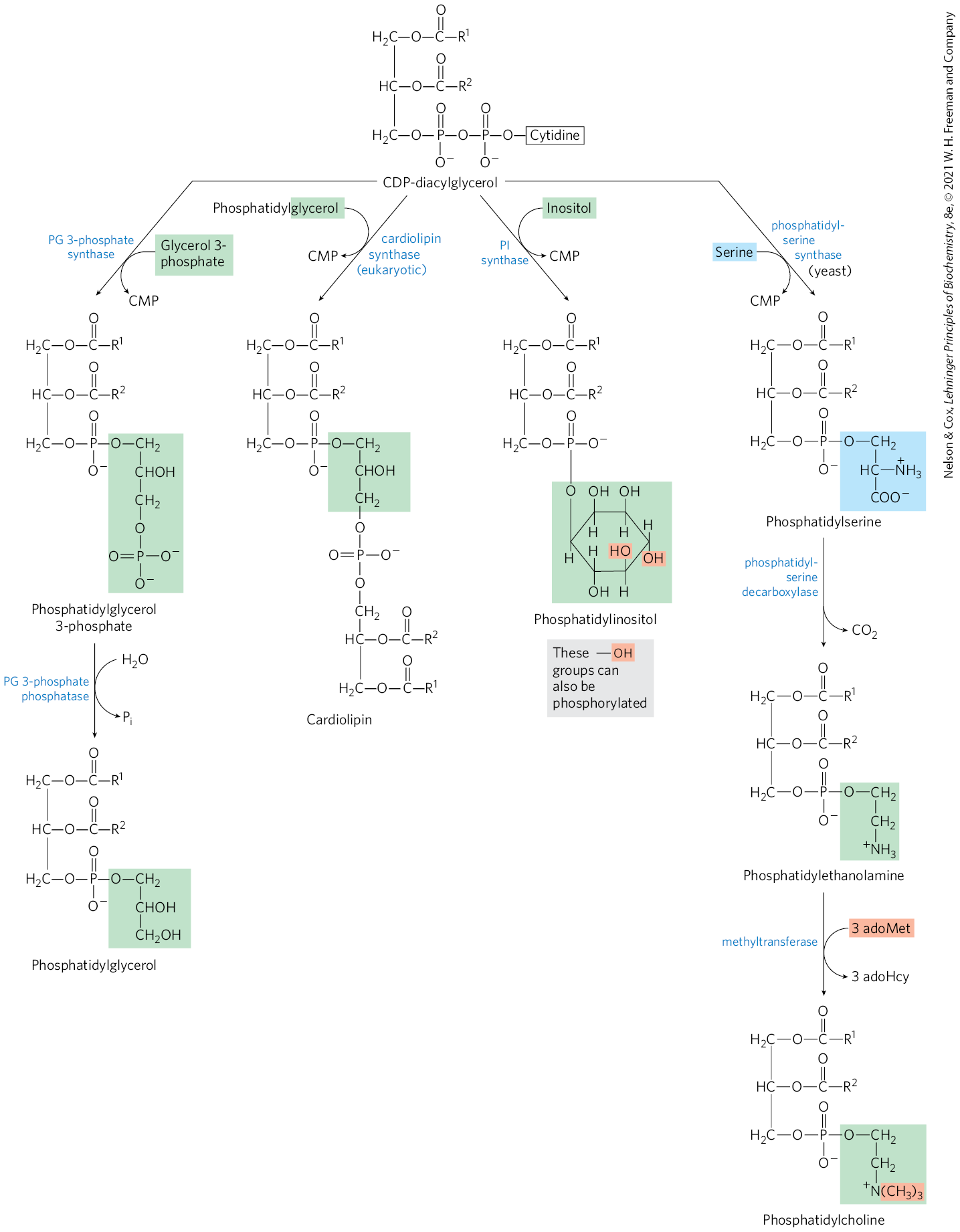
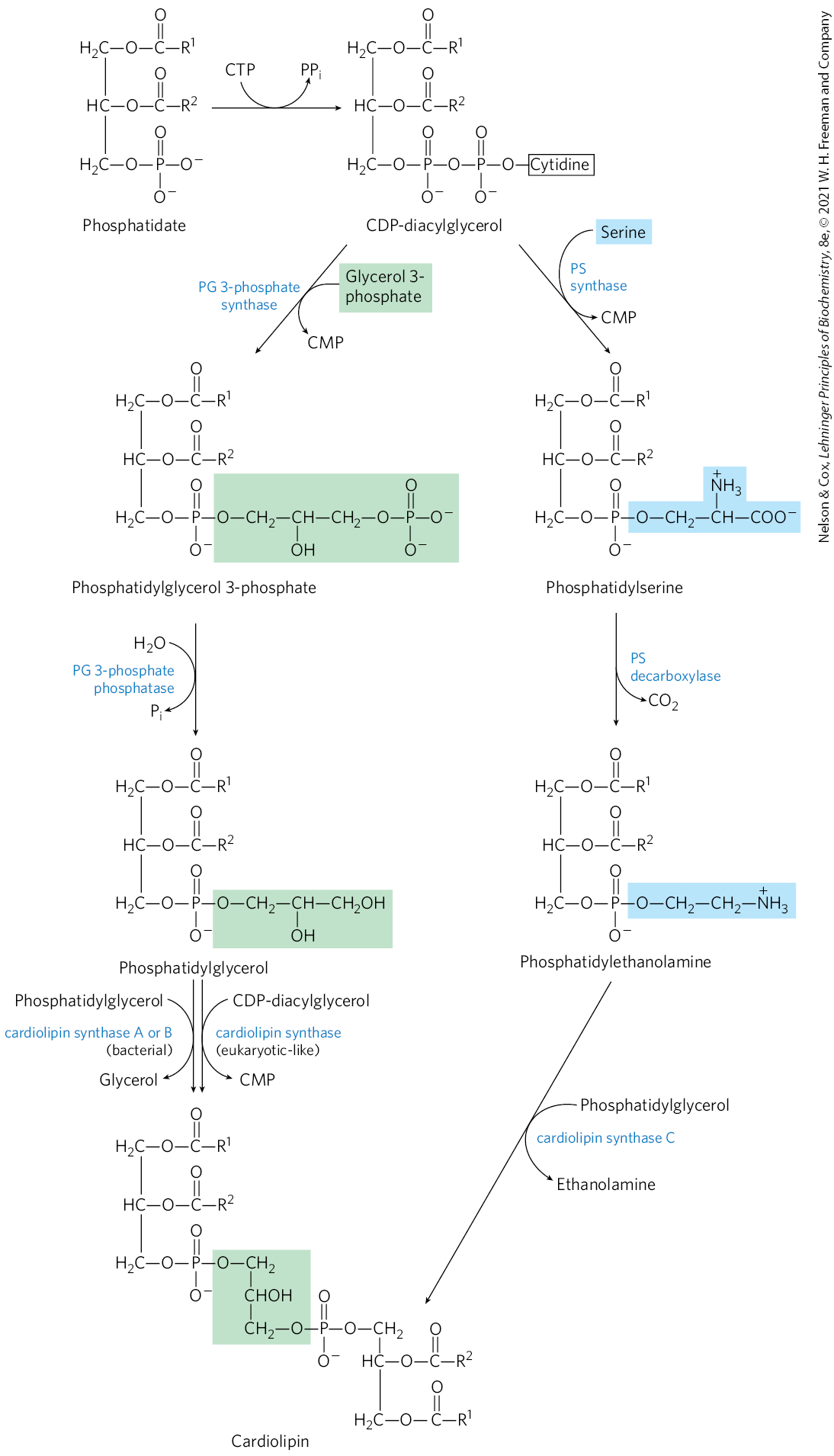
In eukaryotes, many phospholipids are synthesized using strategy 1 in Figure 21-24, and many of the pathways begin with CDP-diacylglycerol. The synthesis of phosphatidylglycerol provides our first example. Beginning with CDP-diacylglycerol (Fig. 21-25), displacement of CMP through nucleophilic attack by the C-1 hydroxyl of glycerol 3-phosphate yields phosphatidylglycerol 3-phosphate. Phosphatidylglycerol 3-phosphate is processed further by cleavage of the phosphate monoester (with release of ) to yield phosphatidylglycerol.
FIGURE 21-25 Synthesis of glycerophospholipids in eukaryotes using CDP-diacylglycerol. These glycerophospholipids are synthesized using strategy 1 in Figure 21-24. Phosphatidylglycerol is synthesized by reaction of CDP-diacylglycerol with glycerol-3-phosphate, followed by dephosphorylation. Phosphatidylglycerol can react with CDP-diacylglycerol to generate cardiolipin. Phosphatidylinositol is generated from CDP-diacylglycerol in a single step. The pathway from CDP-diacylglycerol to phosphatidylserine is used in yeast but not in mammals. The pathway from phosphatidylserine to phosphatidylethanolamine and phosphatidylcholine is common to all eukaryotes.
In eukaryotes, cardiolipin is a relatively uncommon phospholipid, found almost exclusively in the inner membranes of mitochondria. As described below, cardiolipin is important in bacteria, and its presence in mitochondria is likely yet another relic of the bacterial origin of these organelles. Cardiolipin is essential for the function of some mitochondrial enzymes. It is also synthesized using strategy 1, by condensation of CDP-diacylglycerol with phosphatidylglycerol (Fig. 21-25).
Phosphatidylinositol is similarly synthesized by condensation of CDP-diacylglycerol with inositol (Fig. 21-25). Specific phosphatidylinositol kinases then convert phosphatidylinositol to its phosphorylated derivatives. Phosphatidylinositol and its phosphorylated products in the plasma membrane play a central role in signal transduction in eukaryotes (see Figs. 12-11, 12-15, 12-23).
Yeast (but not mammals) use a similar path to produce phosphatidylserine by condensation of CDP-diacylglycerol and serine, and they can synthesize phosphatidylethanolamine from phosphatidylserine in the reaction catalyzed by phosphatidylserine decarboxylase (Fig. 21-25). This pathway for synthesis of phosphatidylethanolamine occurs primarily in the mitochondria, although this lipid is transported from there to other cellular membranes. Phosphatidylethanolamine may be converted to phosphatidylcholine (lecithin) by the addition of three methyl groups to its amino group; S-adenosylmethionine is the methyl group donor (see Fig. 18-18) for all three methylation reactions.
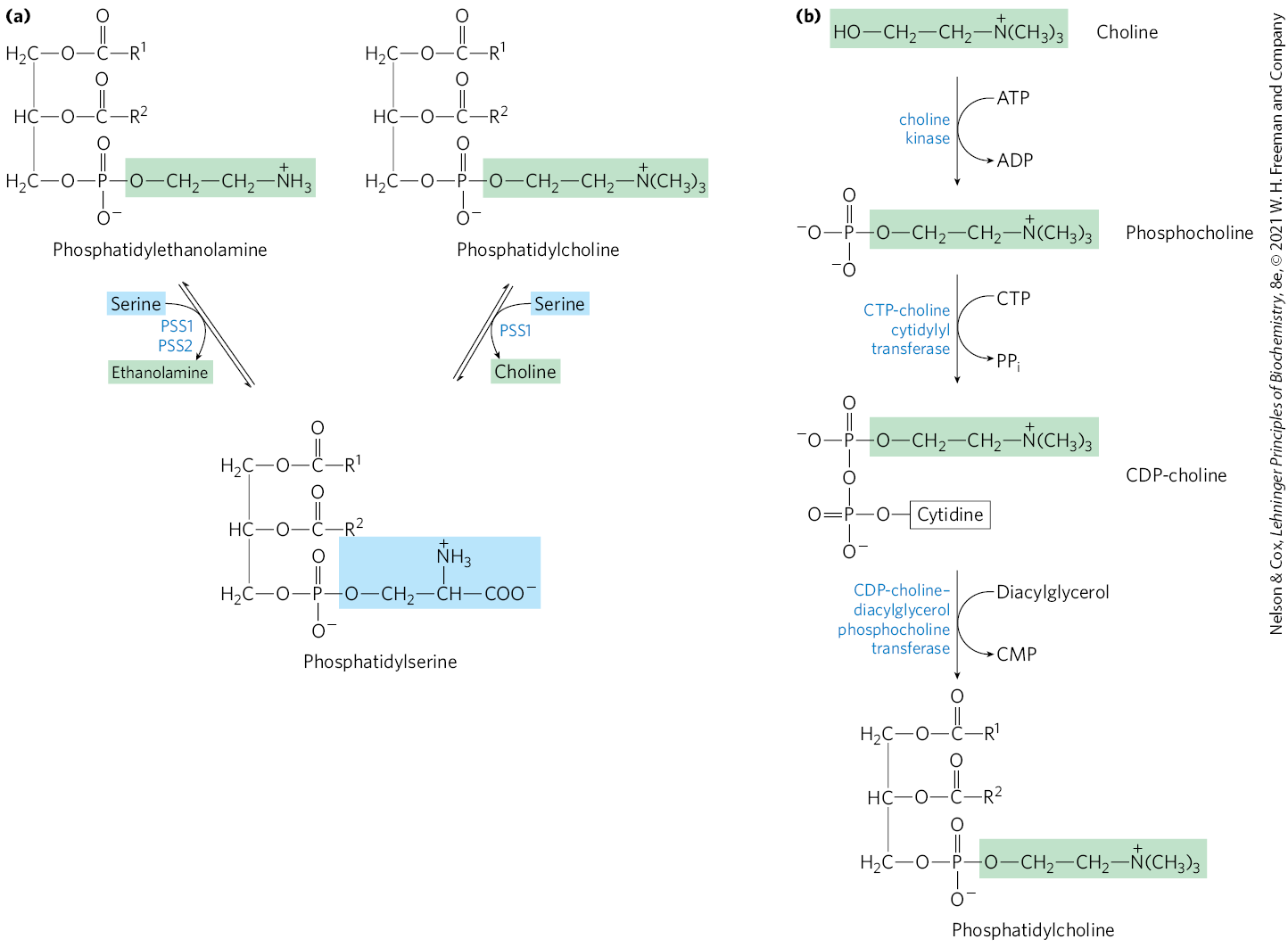
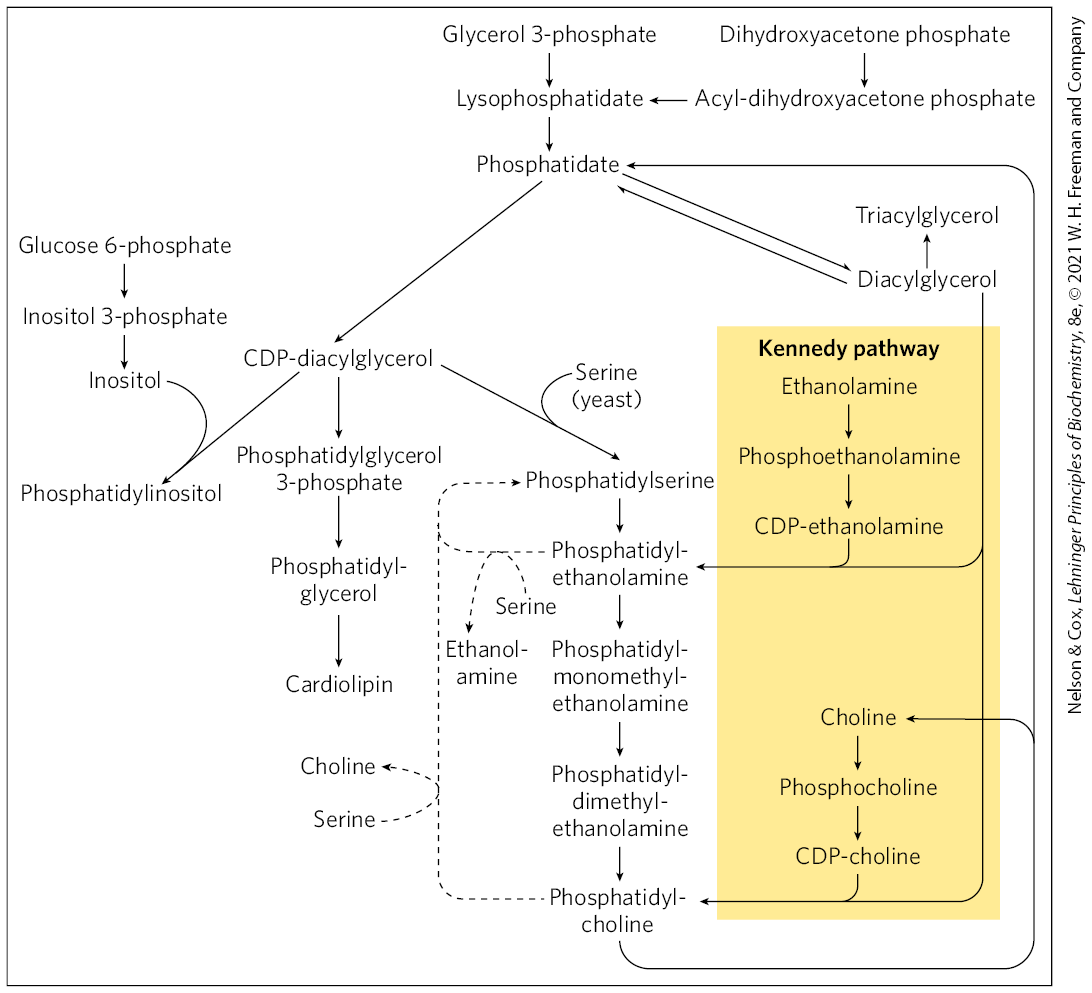
In mammals, strategy 2 (Fig. 21-24) is utilized for the synthesis of phosphatidylethanolamine and phosphatidylcholine in the membranes of the ER and nucleus. The activation of the head group to the CDP derivative is followed by condensation with diacylglycerol as shown for phosphatidylcholine (Fig. 21-26a). These pathways serve to salvage free ethanolamine and choline. In contrast, mammalian phosphatidylserine biosynthesis does not utilize either strategy shown in Figure 21-24; instead, it is derived from phosphatidylethanolamine or phosphatidylcholine via one of two head-group exchange reactions carried out in the ER (Fig. 21-26b). These reactions generate free ethanolamine and choline, respectively. The major sources of phosphatidylethanolamine and phosphatidylcholine in all eukaryotic cells are summarized in Figure 21-27.
FIGURE 21-26 Pathways for phosphatidylserine and phosphatidylcholine synthesis in mammals. (a) Phosphatidylserine is synthesized by -dependent head-group exchange reactions promoted by phosphatidylserine synthase 1 (PSS1) or phosphatidylserine synthase 2 (PSS2). PSS1 can use either phosphatidylethanolamine or phosphatidylcholine as a substrate. (b) The same strategy shown here for phosphatidylcholine synthesis (strategy 2 in Fig. 21-24) is also used for salvaging ethanolamine in phosphatidylethanolamine synthesis.
FIGURE 21-27 Summary of the pathways for synthesis of major phospholipids and triacylglycerols in eukaryotes. Phosphatidic acid is formed by transacylation of l-glycerol 3-phosphate with two fatty acyl groups donated from fatty acyl–CoA. The enzyme phosphatidic acid phosphatase (lipin) converts phosphatidic acid to diacylglycerol, which in the Kennedy pathway condenses with a CDP-activated head group (ethanolamine or choline) to form phosphatidylethanolamine or phosphatidylcholine. Alternatively, phosphatidic acid can be activated with a CDP moiety, which is displaced by condensation with a head-group alcohol — inositol, glycerol 3-phosphate, or serine, forming phosphatidylinositol, phosphatidylglycerol, or (only in yeast and fungi) phosphatidylserine. Decarboxylation of phosphatidylserine yields phosphatidylethanolamine, and methylation of phosphatidylethanolamine produces phosphatidylcholine. In mammals, phosphatidylserine and phosphatidylcholine are generated via the head-group exchange pathways, detailed in Fig. 21-26. Lysophosphatidic acid is phosphatidic acid missing one of the two fatty acyl groups. [Information from G. M. Carman and G.-S. Han, Annu. Rev. Biochem. 80:859, 2011, Fig. 2.]
The most prominent phospholipids in bacteria are phosphatidylglycerol, phosphatidylethanolamine, and cardiolipin. The pathway for phosphatidylglycerol synthesis (Fig. 21-28) is identical to the path employed in mammals (compare to Fig. 21-25), beginning with CDP-diacylglycerol and using strategy 1. Phosphatidylethanolamine is produced in a similar pathway, with phosphatidylserine an intermediate. In bacteria, there are multiple biosynthetic paths to the third prominent phospholipid, cardiolipin, in which two diacylglycerols are joined through a common head group (Fig. 21-28).
FIGURE 21-28 Origin of the polar head groups of phospholipids in E. coli. Initially, a head group (either serine or glycerol 3-phosphate) is attached via a CDP-diacylglycerol intermediate (strategy 1 in Fig. 21-24). For phospholipids other than phosphatidylserine, the head group is further modified, as shown here. In the enzyme names, PG represents phosphatidylglycerol; PS, phosphatidylserine. Cardiolipin can be generated from either phosphatidylglycerol or phosphatidylethanolamine via multiple pathways, as shown. One of these pathways, in which phosphatidylglycerol is condensed with CDP-diacylglycerol, is identical to the pathway used in eukaryotes.
Eukaryotic Membrane Phospholipids Are Subject to Remodeling
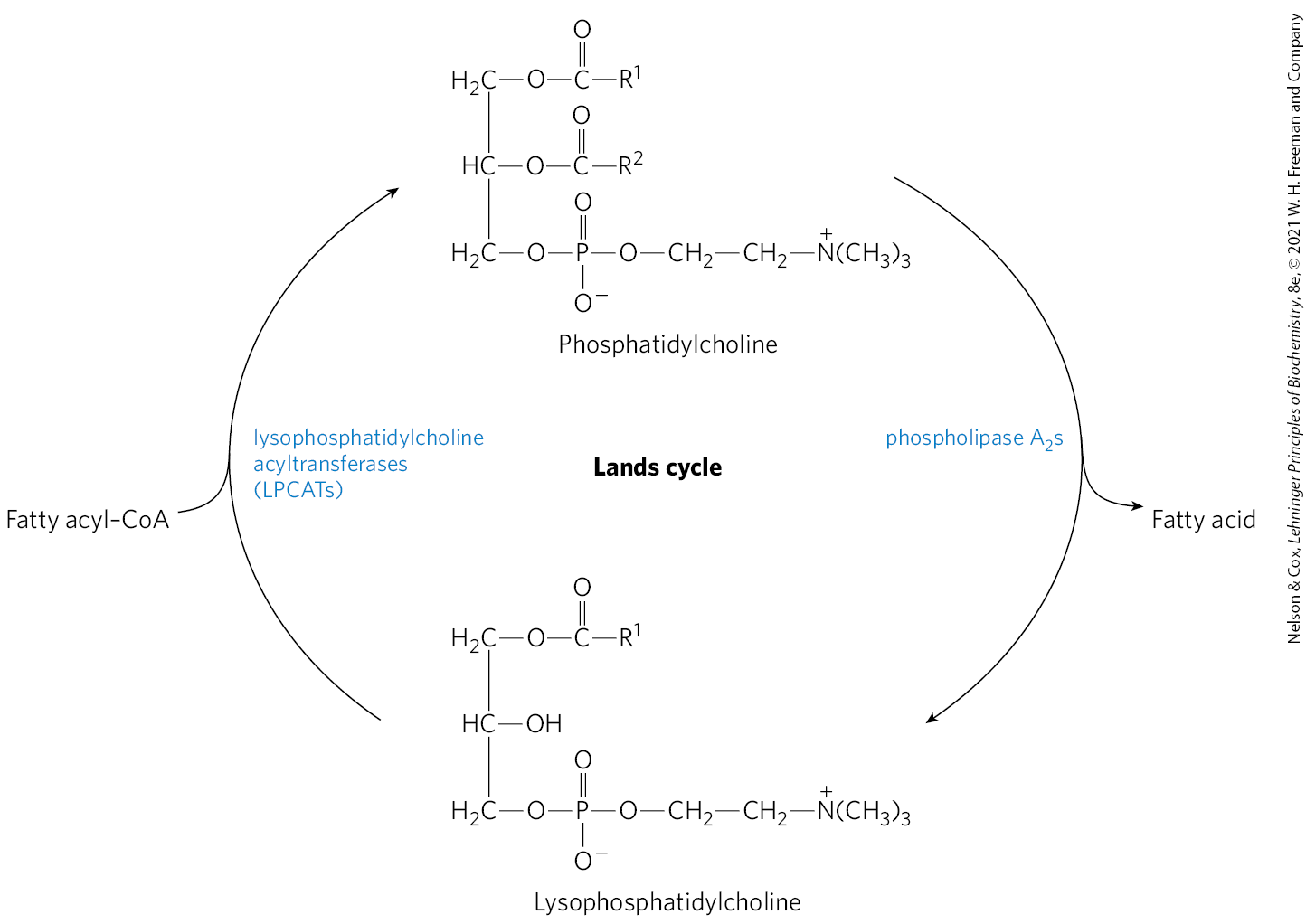
In principle, the two fatty acyl groups esterified to C-1 and C-2 in a phospholipid may vary in length and degree of desaturation, thus altering the properties of the membrane of which it is a part. Phosphatidylcholine is the major structural phospholipid of mammalian membranes, often representing 40% to 50% of the total. Thus, much of the remodeling centers on phosphatidylcholine. The remodeling occurs largely by a process called the Lands cycle (Fig. 21-29), which replaces the polyunsaturated fatty acyl group at C-2. The fatty acyl moieties are first hydrolyzed by phospholipase (Fig. 10-14) to generate 1-acyl lysophospholipids. The fatty acid is then replaced by a class of enzymes called lysophosphatidylcholine acyltransferases, or LPCATs. There are at least four LPCATs in humans, each with distinct tissue distributions and substrate specificities.
FIGURE 21-29 The Lands cycle for phospholipid remodeling. Phosphatidylcholine, which is the most common phospholipid of eukaryotic membranes, is a major target of this process. Fatty acids at C-2 are removed by phospholipase enzymes. A new fatty acid is then introduced by the action of a lysophosphatidylphocholine acyltransferase (LPCAT). The four mammalian LPCATs differ in tissue distribution and substrate specificity.
The physiological effects of LPCAT enzymes go far beyond the alteration of the lipid composition of membranes. LPCAT3, the most widely distributed version of the enzyme, helps to regulate lipogenesis and secretion of very-low-density lipoproteins (VLDLs), described later in this chapter. Mice lacking LPCAT3 have a greatly reduced intake of fatty acids in the intestine, resulting in the release of gut hormones that control appetite. They are unable to survive on a high-fat diet, they resist eating, and they die of starvation unless the diet is changed. LPCATs play a demonstrable but still often mysterious role in processes from atherosclerosis to obesity to cancer, making these enzymes the subjects of increasing interest as research and drug targets.
Plasmalogen Synthesis Requires Formation of an Ether-Linked Fatty Alcohol
The biosynthetic pathway to ether lipids, including plasmalogens and the platelet-activating factor (see Fig. 10-9), requires displacement of an esterified fatty acyl group by a long-chain alcohol to form the ether linkage (Fig. 21-30). Head-group attachment follows, by mechanisms essentially like those used in formation of the common ester-linked phospholipids. Finally, the characteristic double bond of plasmalogens is introduced by the action of a mixed-function oxidase similar to that responsible for desaturation of fatty acids (Fig. 21-13). The peroxisome is the primary site of plasmalogen synthesis.
FIGURE 21-30 Synthesis of ether lipids and plasmalogens. The newly formed ether linkage is shaded light red. The intermediate 1-alkyl-2-acylglycerol 3-phosphate is the ether analog of phosphatidic acid. Mechanisms for attaching head groups to ether lipids are essentially the same as for their ester-linked analogs. The characteristic double bond of plasmalogens (shaded blue) is introduced in a final step by a mixed-function oxidase system similar to fatty acyl–CoA desaturase.
Sphingolipid and Glycerophospholipid Synthesis Share Precursors and Some Mechanisms
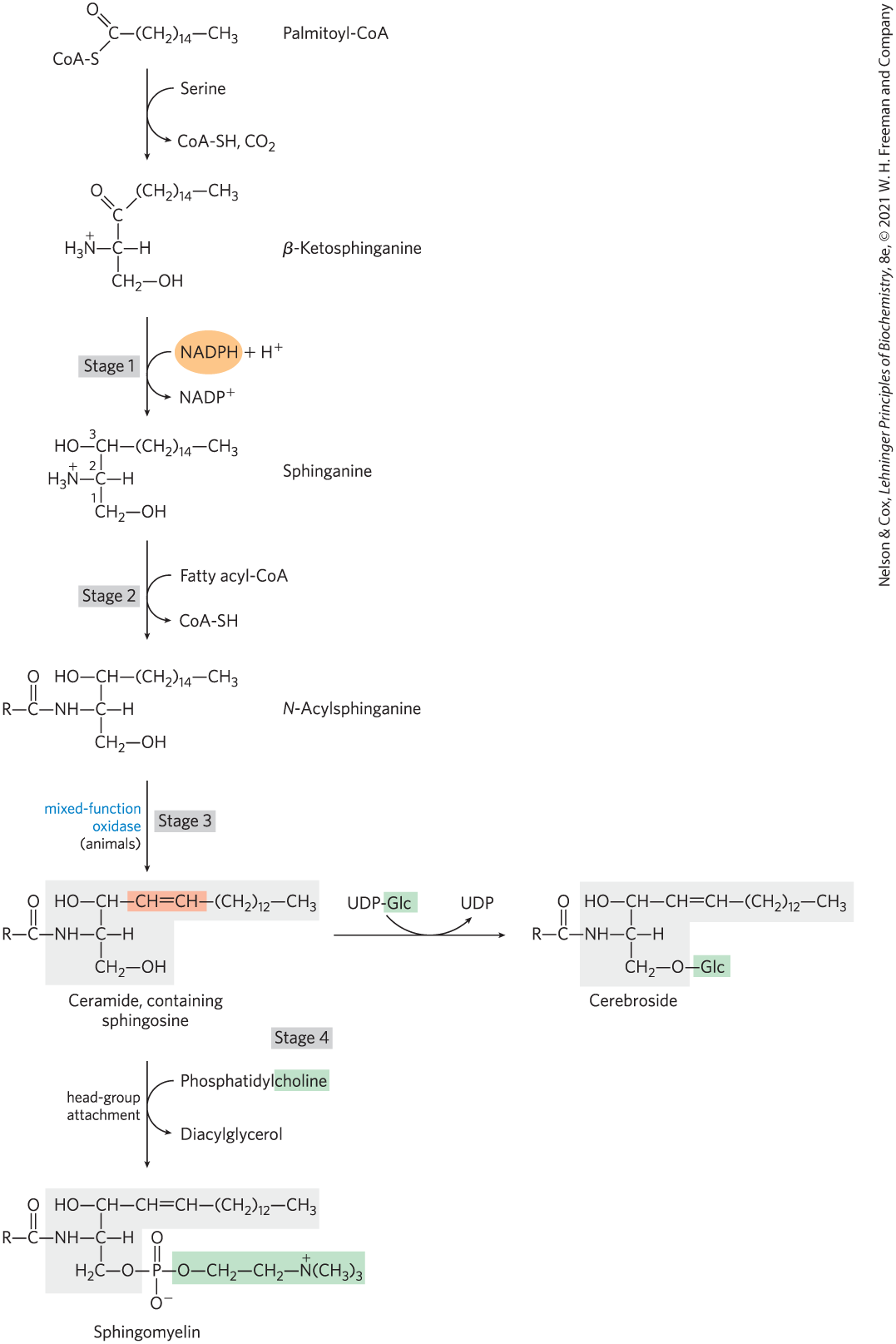
The biosynthesis of sphingolipids takes place in four stages: (1) synthesis of the 18-carbon amine sphinganine from palmitoyl-CoA and serine; (2) attachment of a fatty acid in amide linkage to yield N-acylsphinganine; (3) desaturation of the sphinganine moiety to form N-acylsphingosine (ceramide); and (4) attachment of a head group to produce a sphingolipid such as a cerebroside or sphingomyelin (Fig. 21-31). The first few steps of this pathway occur in the ER; the attachment of head groups in stage 4 occurs in the Golgi complex. The pathway shares several features with the pathways leading to glycerophospholipids: NADPH provides reducing power, and fatty acids enter as their activated CoA derivatives. In cerebroside formation, sugars enter as their activated nucleotide derivatives. Head-group attachment in sphingolipid synthesis has several novel aspects. For example, phosphatidylcholine, rather than CDP-choline, serves as the donor of phosphocholine in the synthesis of sphingomyelin.
FIGURE 21-31 Biosynthesis of sphingolipids. Condensation of palmitoyl-CoA and serine, forming β-ketosphinganine, followed by reduction with NADPH, yields sphinganine, which is then acylated to N-acylsphinganine (a ceramide). The sphingosine is shaded gray. In animals, a double bond (shaded light red) is created by a mixed-function oxidase before the final addition of a head group: phosphatidylcholine, to form sphingomyelin, or glucose, to form a cerebroside.
In glycolipids — the cerebrosides and gangliosides (see Fig. 10-11) — the head-group sugar is attached directly to the C-1 hydroxyl of sphingosine in glycosidic linkage rather than through a phosphodiester bond. The sugar donor is a UDP-sugar (UDP-glucose or UDP-galactose).
Polar Lipids Are Targeted to Specific Cellular Membranes
Membrane lipids are insoluble in water, so they cannot simply diffuse from their point of synthesis (the ER) to their point of insertion. Instead, they are transported from the ER to the Golgi complex, where additional synthesis can take place. They are then delivered in membrane vesicles that bud from the Golgi complex and then move to and fuse with the target membrane (see Fig. 11-4). Sphingolipid transfer proteins carry ceramide from the ER to the Golgi complex, where sphingomyelin synthesis occurs. Cytosolic proteins also bind phospholipids and sterols and transport them between cellular membranes (see Fig. 11-7). These mechanisms contribute to establishment of the characteristic lipid compositions of organelle membranes (see Fig. 11-5).
SUMMARY 21.3 Biosynthesis of Membrane Phospholipids
Beginning with diacylglycerol precursors, there are two pathways for adding head groups to phospholipids. Either the diacylglycerol (strategy 1) or the head group (strategy 2) is activated by CDP.
In eukaryotes, phospholipid biosynthetic strategies vary with subcellular location. The major phospholipids phosphatidylethanolamine and phosphatidylcholine are synthesized using strategy 1 in the mitochondria and strategy 2 in the ER and nucleus. Phosphatidylserine is derived from head-group exchange with phosphatidylethanolamine or phosphatidylcholine.
In mammals, phospholipids are remodeled in membranes via the Lands cycle. Remodeling is facilitated by lysophosphatidylcholine acyltransferases.
The characteristic double bond in plasmalogens is introduced by a mixed-function oxidase.
The head groups of sphingolipids are attached by unique mechanisms.
Phospholipids travel to their intracellular destinations via transport vesicles or specific proteins.

 The physiological effects of LPCAT enzymes go far beyond the alteration of the lipid composition of membranes. LPCAT3, the most widely distributed version of the enzyme, helps to regulate lipogenesis and secretion of very-low-density lipoproteins (VLDLs), described later in this chapter. Mice lacking LPCAT3 have a greatly reduced intake of fatty acids in the intestine, resulting in the release of gut hormones that control appetite. They are unable to survive on a high-fat diet, they resist eating, and they die of starvation unless the diet is changed. LPCATs play a demonstrable but still often mysterious role in processes from atherosclerosis to obesity to cancer, making these enzymes the subjects of increasing interest as research and drug targets.
The physiological effects of LPCAT enzymes go far beyond the alteration of the lipid composition of membranes. LPCAT3, the most widely distributed version of the enzyme, helps to regulate lipogenesis and secretion of very-low-density lipoproteins (VLDLs), described later in this chapter. Mice lacking LPCAT3 have a greatly reduced intake of fatty acids in the intestine, resulting in the release of gut hormones that control appetite. They are unable to survive on a high-fat diet, they resist eating, and they die of starvation unless the diet is changed. LPCATs play a demonstrable but still often mysterious role in processes from atherosclerosis to obesity to cancer, making these enzymes the subjects of increasing interest as research and drug targets. 
 The biosynthetic pathway to ether lipids, including
The biosynthetic pathway to ether lipids, including 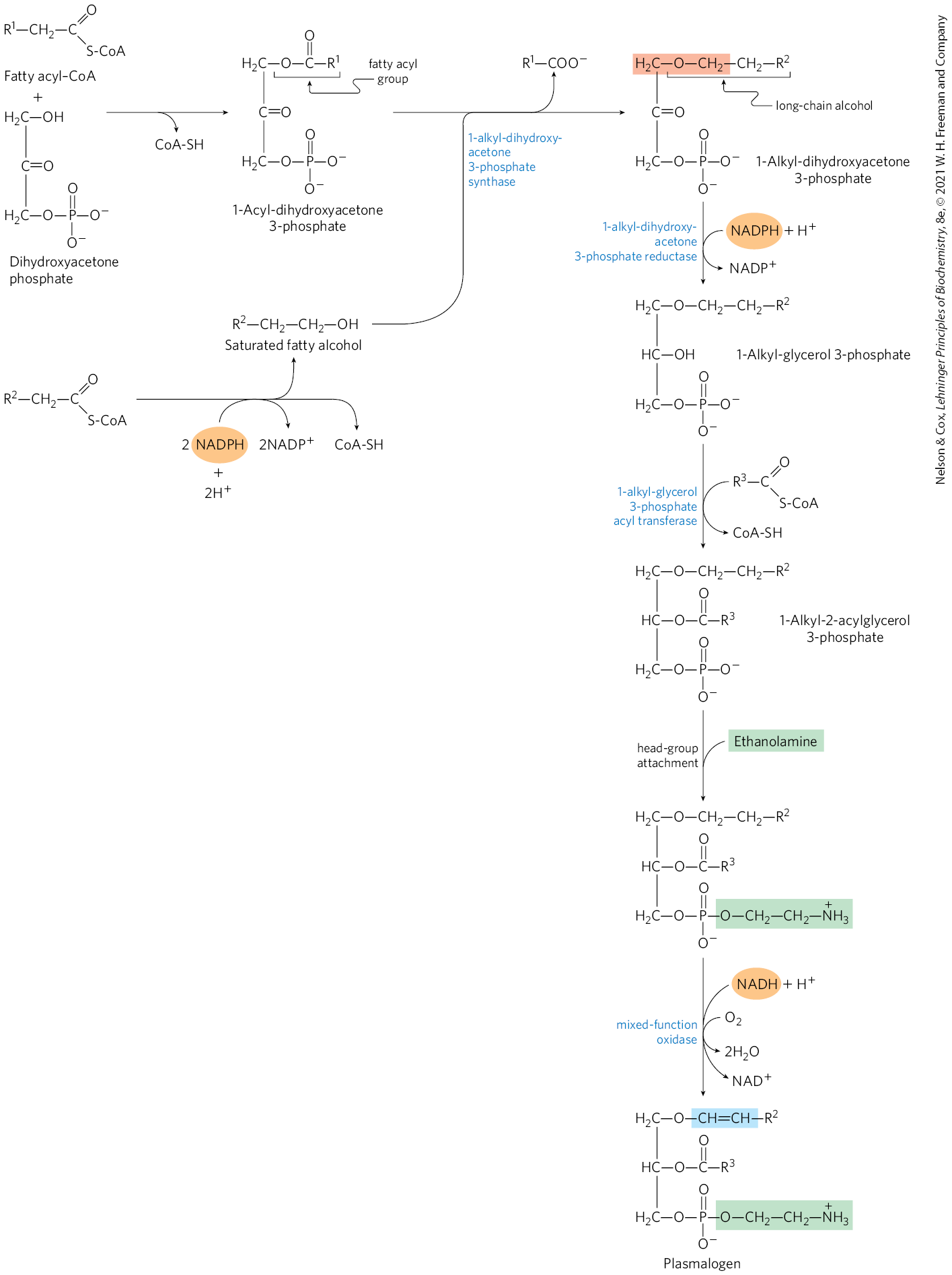
 Beginning with diacylglycerol precursors, there are two pathways for adding head groups to phospholipids. Either the diacylglycerol (strategy 1) or the head group (strategy 2) is activated by CDP.
Beginning with diacylglycerol precursors, there are two pathways for adding head groups to phospholipids. Either the diacylglycerol (strategy 1) or the head group (strategy 2) is activated by CDP.